The Origin of The Iberomaurusians
Chapters:
The Jebel Irhoud Fossils

The oldest known human inhabitants of North Africa are represented by fossils that were discovered in Jebel Irhoud - Morocco [Link]; these fossils have been estimated to be 286,000 to 315,000 years old. The morphology of these fossils appears to be intermediate between Homo heidelbergensis and Homo sapiens, given the theory that the former evolved into the latter. For example, Jebel Irhoud 1 has large brow ridges and a cranium that's elongated along its saggital plane, which are traits of Homo heidelbergensis; however, it lacks the occipital bun of Homo heidelbergensis and its cranium is positioned above its face due to having a vertical frontal bone, which are traits of Homo sapiens.

Due to reasons such as these, the Jebel-Irhoud humans were likely basal Homo sapiens - meaning that they were at the beginning of the transition from Homo heidelbergensis to Homo sapiens.
The consensus of anthropologists is that Homo erectus evolved into Homo heidelbergensis, and Homo heidelbergensis evolved into Homo sapiens. Considering that the oldest fossils of Homo erectus, Homo heidelbergensis, Homo sapiens idaltu, and AMHs (anatomically modern humans, the only remaining subspecies of Homo sapiens and the only remaining member of Homo) have been found in East Africa, the lineage that is comprised of these species and subspecies must have originated and developed in East Africa. Therefore, if the Jebel-Irhoud humans were indeed at the beginning of the transition from Homo heidelbergensis to Homo sapiens, they must have belonged to this lineage, which means that they must have come from East Africa.
However, they must have left the lineage before it produced AMHs or Homo sapiens idaltu, since they inhabited Northwest Africa rather than East Africa. Therefore, they either died off without producing any descendants … or they evolved into another subspecies of Homo sapiens.

Fossils of each species in the evolutionary lineage that produced Homo sapiens, starting with Homo erectus and ending with AMHs (anatomically modern humans), who are the only remaining subspecies of Homo sapiens and the only remaining member of the genus Homo.
Note that all of these fossils - except for Jebel Irhoud 1 - were discovered in East Africa. This indicates that East Africa is where the lineage that produced Homo sapiens evolved and that the Jebel Irhoud humans were migrants from East Africa.
The Aterians

A Reconstruction of the Aterian fossil Dar-Es Sultane II H5 by Ancestral Whispers [Link]
The second oldest known human inhabitants of North Africa were the Aterians [Link], whose stone-tool culture flourished therein from 150,000 YBP to 20,000 YBP, according to archaeological evidence. Archaeological sites of their stone-tool culture are located in Morocco, Algeria, Tunisia, Niger, Libya, and Egypt.

Map Highlighting Archeological Sites of the Aterian Stone Tool Culture [Link]
The Aterians are represented by the human fossils of Dar-Es Sultane - Morocco, which have been determined to be 100,000 years old. Their morphology shares continuity with that of the Jebel-Irhoud human fossils and they were found in the same country as them, which indicates that the former evolved into the latter. Their morphology is similar to that of AMHs in some ways but is different in other ways, suggesting that they may have been the same species as AMHs - but they may have also been different enough to be a separate subspecies.

Photo of Dar-Es Sultane II H5 (left) and Jebel Irhoud 1 (right)
The following is a quote from “The origin and Evolution of Homo sapiens” (2016) [Link] about the Aterian human fossil Dar-Es Sultane II H5. Note that the quote likens it to both one of the Jebel-Irhoud human fossils and fossils of AMHs that were discovered in Qafzeh - Israel, which supports the idea that the Aterians were both different from AMHs but also similar - as a fellow subspecies would be.
“Deciding on how to classify Dar-Es Sultane II H5 is difficult. It has a rather modern looking face and frontal bone shape, but both are very large in size, as is the supraorbital development. Although previous assessments have suggested that it could represent an Aterian intermediate between the Middle Stone Age associated Irhoud specimens and those of the Iberomaurusian, (i.e. the local late Upper Paleolithic) morphometric analyses place it closer to Jebel Irhoud 1 and the Qafzeh crania than to the late Pleistocene fossils.”
Due to the likelihood that the Aterians descended from the Jebel-Irhoud humans, who were very likely basal Homo sapiens, and due to being both similar and different relative to AMHs, a conclusion that is being drawn herein is that the Aterians were indeed a separate subspecies of Homo sapiens.

Skhul V and Qafzeh 9 and reconstructions of them by Kennis & Kennis [Link]
In addition to being a fellow subspecies of Homo sapiens, another reason that Dar-Es Sultane II H5 bears morphometric proximity to the AMH (anatomically modern human) Qafzeh crania could be interbreeding. It is not unusual for breeding to occur between different subspecies - or even different species of the same genus - when they occasionally encounter one another. Therefore, DNA of AMHs could have introgressed into the lineage that was comprised of the Jebel-Irhoud Humans and the Aterians.
This possibility is supported by the age of the Qafzeh fossils and the location at which they were discovered: Qafzeh - Israel, as well as the age of another set of AMH fossils and the fact that they were also discovered in Israel: the Skhul fossils [Link]. Both sets of fossils have collectively been estimated to be 80,000 to 120,000 years old, which makes them younger than the fossils of AMHs that were discovered in East Africa: those of Omo Valley - Ethiopia, which have been estimated to be 233,000 years old and are therefore the oldest.
The ages and the locations at which these fossils were discovered imply that AMHs were migrating from East Africa - where they evolved - into Israel, which means that they would have traveled through North Africa (through Egypt) in order to enter Israel. Therefore, it is likely that some of these individuals remained in North Africa and hybridized with members of the lineage that was comprised of the Jebel-Irhoud Humans and the Aterians.
The Iberomaurusians

A Reconstruction of an Iberomaurusian man by Ancestral Whispers [Link]
Origin
The third oldest known human inhabitants of North Africa are the Iberomaurusians [Link], whose culture has been estimated to have appeared therein approximately 25,000 YBP. This date was determined via radio-carbon dating of organic remains that were discovered at Tamar Hat - Algeria, which is one of several archaeological sites of the Iberomaurusians; this is explained in the peer-reviewed scientific article “New radiocarbon dates for the earliest Later Stone Age microlithic technology in Northwest Africa” (2016) [Link].
“The new radiocarbon ages for Tamar Hat are closely comparable to the ages originally obtained by Saxon et al. (1974), but are more reliable because they are on bone collagen which has fewer issues with possible contamination. The calibrated ages indicate a timespan of at least 5000 years for the LSA [Later Stone Age] occupation, beginning near the base at 25,845–25,270 cal BP and ending at around 20,122–19,632 cal BP. It is noticeable that the industry appears ‘suddenly’ and with no obvious antecedence.”
This states that Tamar Hat was occupied from possibly as early as 25,845 YBP, which is the oldest time period to which any Iberomaurusian site has been dated. Therefore, this date may have been when the Iberomaurusian culture appeared in North Africa.
Additional evidence of this date are the maternal lineages of Iberomaurusian human fossils that were discovered at Taforalt - Morocco; they possess Maternal Haplogroups U6a and M1b, which have been estimated to have formed approximately 25,000 YBP, which was close to the aforementioned date of 25,845 YBP. This is explained in the following quote from the peer-reviewed scientific article “Pleistocene North African Genomes Link Near Eastern And Sub-Saharan African Human Populations” (2018) [Link].
“Here we present genome-wide data from seven individuals, directly dated between 15,100 and 13,900 calibrated years before present (cal. yr B.P.), from Grotte des Pigeons near Taforalt in eastern Morocco…. Mitochondrial consensus sequences of the Taforalt individuals belong to the U6a (six individuals) and M1b (one individual) haplogroups, which are mostly confined to present-day populations in North and East Africa.… Notably, the diversification of haplogroups U6a and M1 found for Taforalt is dated to ~24,000 yr B.P., which is close in time to the earliest known appearance of the Iberomaurusian culture in Northwest Africa [25,845 to 25,270 cal. yr B.P. at Tamar Hat].”
As for where the Iberomaurusians came from, morphological continuity between the Aterians and the Iberomaurusians strongly indicates that the latter descended from the former. This is stated in a quote from the peer-reviewed scientific article “The relevance of late MSA mandibles on the emergence of modern morphology in Northern Africa” (2022) [Link]. Note that MSA means Middle Stone Age.
“In Northern Africa, Aterians display similarities to Iberomaurusians and recent humans in the area as well as to the Tighenif and Thomas Quarry hominins, suggesting a greater time depth for regional continuity than previously assumed…. Iberomaurusians are clearly larger than their contemporaries, covering a similar size range as Aterians and early H. sapiens…. Even though we have no proof of an in-situ population succession, Aterian morphology fits the human fossil gap between Jebel Irhoud 11 and Iberomaurusians, suggesting a greater time depth for regional continuity in Northern Africa than previously established.”

Figure 7 from “The relevance of late MSA mandibles on the emergence of modern morphology in Northern Africa” (2022) [Link], which depicts virtual reconstructions of human mandibles. The mandible that is depicted at the bottom and that is colored turquoise is Taforalt XVIII, which is an Iberomaurusian specimen; superimposed onto it is the mandible of El Harhoura, which is an Aterian specimen. Aterians had mandibles with wider rami (the rear, vertical components) and taller main bodies relative to those of AMHs, and these traits are shared by the Iberomaurusian specimen. This is consistent with the possibility that the Iberomaurusians descended from the Aterians.
Due to these morphological similarities, the successive time periods of the Aterian and Iberomaurusian cultures, and their presence in the same region (i.e. North Africa), a conclusion that is being drawn herein is that the Iberomaurusians did indeed descend from the Aterians. However, the Iberomaurusians could not have descended from the Aterians exclusively; they possessed autosomal, maternal and paternal DNA of AMHs, which means that they were hybrids (since it has been concluded herein that the Aterians were a different subspecies of Homo sapiens relative to AMHs).
The Source of the Iberomaurusians’ Maternal Haplogroups
The terms sub-Saharan African and SSA refer to people whose ancestors never migrated out of Africa; alternatively, they refer to the ancestry of such people or to DNA that is associated with it. Note that sub-Saharan African & SSA refer to multiple lineages that diverged from one another tens of thousands to over two-hundred thousand years ago; therefore, these lineages are only distantly related to each other.
The term Eurasian refers to people who descend from migrants who left Africa tens of millennia ago and settled in Eurasia - and whose subsequent lineages experienced genetic drift and evolved adaptations to environments therein; alternatively, it refers to the ancestry of such people or to DNA that is associated with it.
Oceanians and Indigenous Americans descend from Eurasians. Therefore, they can be thought of as part of the Eurasian clade.
________________________
All maternal haplogroups that are labeled with the letter “L” are sub-Saharan African. Conversely, all maternal haplogroups that are labeled with a letter other than “L” are Eurasian.
However, it must be noted that all Eurasian maternal haplogroups descend from L3, which originated in sub-Saharan Africa and which has many subclades that are exclusive to sub-Saharan Africans; in other words, Eurasian maternal haplogroups are a subset of sub-Saharan Africa’s L3, but they are demarcated in such a way that reflects their association with Eurasians. Therefore, any non-L maternal haplogroup that is present in Africa was brought to the continent via a back-migration, or it descends from one that was brought into the continent via a back-migration.
As already stated, the Iberomaurusian human fossils of Taforalt (who will be referred to as the “Taforalt specimens” from hereon) possess Maternal Haplogroups U6a and M1b, which directly or indirectly come from West Asia / western Eurasia; here is another quote from the peer-reviewed scientific article “Pleistocene North African Genomes Link Near Eastern And Sub-Saharan African Human Populations” (2018) [Link].
“Mitochondrial consensus sequences of the Taforalt individuals belong to the U6a (six individuals) and M1b (one individual) haplogroups, which are mostly confined to present-day populations in North and East Africa. U6 and M1 have been proposed as markers for autochthonous Maghreb ancestry, which might have been originally introduced into this region by a back-to-Africa migration from West Asia.”
In regard to U6 being present in North Africa as a result of a back-migration, an ancestral form of U6 was discovered in a Romanian human skull that has been named PM1, which means Peștera Muierii 1; it has been estimated to be 35,000 years old and possesses an ancestral form of U6. Therefore, it’s likely that U6 originated in West Asia / western Eurasia.
Quote from the scientific article “Out of Africa – and back again: When did humans return to Africa?” (2016) [Link]:
“… humans might have been migrating back into Africa remarkably early, according to clues found in the mitochondrial DNA of a 35,000-year-old skull discovered in a cave in Romania…. U6 is found in a relatively large proportion of the population in north Africa," he explains. But "this is much older, much more basal, so we demonstrate that the origin of these populations from north Africa were basically from western Eurasia.”
Therefore, a conclusion that is being drawn herein is that Eurasian AMHs were the source of the Taforalt specimens’ maternal haplogroups. As for the identity of these Eurasian AMHs, consider the Natufian component of a two-way admixture model of the Taforalt specimens, which is presented in the aforementioned peer-reviewed scientific article “Pleistocene North African Genomes Link Near Eastern And Sub-Saharan African Human Populations” (2018) [Link].
“A two-way admixture model, comprising Natufian and sub-Saharan African populations, does not significantly deviate from our data, with 63.5% Natufian and 36.5% sub-Saharan African ancestry, on average.”
Note that the use of the Natufians as a component in this model does not necessarily mean that the Taforalt specimens descended from them; it simply means that - at the very least - the Taforalt specimens shared ancestry with the Natufians - and that therefore the Natufians can be used to model this ancestry.
In regard to the SSA component, it obviously does not model the Eurasian source of the Taforalt specimens’ maternal haplogroups, but it will be addressed later herein; instead, the Natufian component is what models the Eurasian source of the Taforalt specimens’ maternal haplogroups.

A Reconstruction of a Natufian man by Ancestral Whispers [Link].
The Natufians [Link] were semi-nomadic hunter-gatherers who may have been the first agriculturalists in the world; their culture flourished within the Levant (e.g. Israel, Jordan, Lebanon, Syria, etc) between 15,000 YBP and 11,500 YBP. However, their culture began ~10,000 years after the Iberomaurusian culture began, and it ended earlier as well.
Iberomaurusian Culture: ~25,000 YBP to ~11,000 YBP
Natufian Culture: 15,000 YBP to 11,500 YBP
Therefore, due to the aforementioned shared ancestry between the Natufians and the Iberomaurusian Taforalt specimens - as well as the later temporal range of the Natufian culture relative to that of the Iberomaurusian culture - a conclusion that is being drawn herein is that the Natufians descended from the Iberomaurusians; in other words, a segment of the Iberomaurusians’ population migrated into the Levant sometime before 15,000 YBP and developed the Natufian culture.
This explains why the Natufians can be used to model a portion of the Taforalt specimens’ DNA. However, it must be noted that they were not genetically identical to the Iberomaurusians, which is why they can be used to model only a portion of the Taforalt specimens’ DNA; this is due to the Natufians having more Arabian and West Asian DNA but less Northern African and sub-Saharan African DNA, as explained in the following quote from the peer-reviewed scientific article, “Re-analysis of Whole Genome Sequence Data From 279 Ancient Eurasians Reveals Substantial Ancestral Heterogeneity” (2018) [Link]:
“The Natufian sample consisted of 61.2% Arabian, 21.2% Northern African, 10.9% Western Asian, and 6.8% Omotic [SSA] ancestry. The transition in the Levant from the Epipaleolithic to the Neolithic period involved an increase of Arabian ancestry at the expense of Northern African and Omotic ancestries.”
Omotic people are sub-Saharan Africans who are indigenous to Ethiopia (e.g. the Aari people [Link]), and ancestry like theirs comprises only 6.8% of the Natufians’ DNA. This means that the Natufians appear to have 29.7% less SSA ancestry than the Taforalt specimens (36.5% - 6.8% = 29.7%). However, the quote also indicates that the Natufians had less Northern African ancestry than their ancestors due to an increase of Arabian ancestry, which implies that there is Northern African ancestry in the Natufian component of the two-way admixture model.
36.5% SSA + 63.5% Natufian = 36.5% SSA + 63.5% ( … + Northern African + … )
This raises the following question: What is Northern African ancestry?
Modern North Africans are partly Eurasian and partly sub-Saharan African, like their Iberomaurusian ancestors. However, the particular combination of Eurasian and SSA ancestry that North Africans have is unique and is therefore defined as Northern African, rather than as individual Eurasian and SSA components of ancestry. This means that there is hidden SSA ancestry in the 21.2% Northern African ancestry of the Natufians.
North Africans are up to 55% sub-Saharan African (55% SSA):
Quote from “Genomic Ancestry of North Africans Supports Back-to-Africa Migrations” (2012) [Link]: “At k = 6 through 8, all North African populations except for Tunisians have sub-Saharan ancestry, present in most individuals, though this ancestry varies between 1%–55%.”
Therefore, there is the following:
21.2% of 55% SSA = 0.212(55% SSA) = 11.66% SSA
By adding this 11.66% SSA to the 6.8% Omotic [SSA] ancestry in the Natufians, their potential total SSA ancestry can be determined:
11.66% SSA + 6.8% SSA = 18.46% SSA
Given that the Natufians must have inherited their SSA ancestry from the Iberomaurusians, the entirety of this 18.46% would be in the Natufian component of the Taforalt specimens’ two-way admixture model. Therefore, there is the following:
36.5% SSA + 18.46% hidden SSA = 54.96% SSA
63.5% Natufian - 18.46% hidden SSA = 45.04% Natufian
Re-expressed two-way admixture model: 54.96% SSA + 45.04% Natufian
Therefore, the actual difference between the amounts of SSA ancestry that the Iberomaurusians and Natufians had is simply the SSA component of the original two-way admixture model:
54.96% SSA - 18.46% SSA = 36.5% SSA
This should not be surprising. Only the SSA ancestry that the Natufians inherited from their Iberomaurusian ancestors should be in the Natufian component; the 36.5% SSA component of the original model is unique to the Taforalt specimens relative to the Natufians and therefore is not in the Natufian component.
The 54.96% SSA component of the re-expressed two-way admixture model will be addressed later herein, as it is verified by another peer-reviewed scientific article - and it is relevant to another subject that will also be addressed later herein.
________________________
The Source of the Iberomaurusians’ Paternal Haplogroup
The Taforalt specimens possess Paternal Haplogroup E-M78 (E1b1b1a1), as stated in yet another quote from the peer-reviewed scientific article “Pleistocene North African Genomes Link Near Eastern And Sub-Saharan African Human Populations” (2018) [Link].
“Consistently, we find that all males with sufficient nuclear DNA preservation carry Y haplogroup E1b1b1a1 (M-78). This haplogroup occurs most frequently in present-day North and East African populations.”
Paternal Haplogroup E-M78 (E1b1b1a1) is a “cousin” of the Natufians’ paternal haplogroup, E-Z830 (E1b1b1b2), which is proof that the Natufians and their Iberomaurusian contemporaries (the Taforalt specimens) descended from a common source. It has been concluded herein that this common source was an older generation of the Iberomaurusians, which means that the Natufians’ split from the Iberomaurusians.
Regarding the Natufians’ paternal haplogroup, the following is a quote from the peer-reviewed scientific article “Placing Ancient DNA Sequences into Reference Phylogenies” (2022) [Link]
“Apart from those described above, the majority (n = 11) of other East African PN samples were placed in E1b1b1b2b-V1515 lineages, a sub lineage of E1b1b1b2-Z830 found in the Levantine proto-agriculturalist Natufians …”
The relation between E-M78 and E-Z830 can be observed via their paths of descent. They are each separated from E-M35 by only one subclade that is upstream relative to themselves, which makes them “cousins.”
Due to E-L539 and E-Z827’s date of formation being 23,900 YBP [Link and Link], neither of them would have existed when the Iberomaurusian culture emerged ~25,000 YBP; this means that E-M35 would have been the most downstream paternal haplogroup of the first Iberomaurusians. Therefore, E-M35 will be referred to from hereon as the paternal haplogroup of the first Iberomaurusians or as the Iberomaurusians’ original paternal haplogroup.
Paternal Haplogroup E-M35 descends from Paternal Haplogroup E-M96, as is shown below:
E-M96 -> E-M5479 -> E-P147 -> E-P177 -> E-P2 (E1b1) -> E-M215 (E1b1b) -> E-M35 [Link]
There is disagreement regarding the origin of E-M96; it may have originated in East Africa according to some sources [Link] - or Eurasia according to other sources [Link]. The disagreement is due to E-M96’s association with Paternal Haplogroups C-M130, F-M89, and D-CTS3946, all three of which are exclusively possessed by Eurasians, Oceanians, and Indigenous Americans - with the exception of D0 [D-Zero].
Regarding Do [D-Zero], it has been detected in a very small number of sub-Saharan Africans. Here is a quote from “A Rare Deep-Rooting D0 [D-Zero] African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa” (2019):
“Here, we reinvestigate a rare deep-rooting African Y-chromosomal lineage by sequencing the whole genomes of three Nigerian men.… We confirm that these three chromosomes do represent a deep-rooting DE lineage, branching close to the DE bifurcation, but place them on the D branch as an outgroup to all other known D chromosomes, and designate the new lineage D0 [D-Zero].”
D0’s [D-Zero’s] presence in sub-Saharan Africa will be addressed later in this section, but first the overall relation between C-M130, F-M89, D-CTS3946, and E-M96 must be explained.
C-M130, F-M89, D-CTS3946, and E-M96 all descend from Paternal Haplogroup CT, as is shown below:
CT -> CF & DE (68,500 YBP)
CF -> C-M130 & F-M89 (65,900 YBP)
DE -> D-CTS3946 & E-M96 (65,200 YBP)
[Link]
All extant paternal haplogroups that formed before CT (e.g. A00, A1b1, etc) - and the paternal haplogroup that is the “brother” of CT (i.e. B-M60) - are carried exclusively by sub-Saharan Africans or people with SSA ancestry; additionally, these paternal haplogroups are more closely related to the precursors of CT (e.g. A-0T, BT, etc) [Link] than those of Eurasians, Oceanians, and Indigenous Americans. Therefore, sub-Saharan Africa is the default location to consider as the birthplace of CT.
Furthermore, as explained in the first section herein, the lineage of species and subspecies to which AMHs belong must have originated in East Africa - due to the oldest fossils of these species and subspecies being discovered therein. Therefore, East Africa is the specific default location to consider in regard to the birthplace of CT, as CT is a paternal haplogroup of AMHs. This is the consensus of anthropologists and geneticists [Link].
However, there is disagreement in regard to the birthplace(s) of the haplogroups that descend from CT, due to the range of estimated dates within which AMHs are theorized to have migrated out of Africa. Some sources say that the migration of AMHs from the Motherland occurred as early as 70,000 YBP, while others say that it occurred as late as 50,000 YBP. Consider the following examples:
“A Rare Deep-Rooting D0 [D-Zero] African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa” (2019) [Link]: “Present-day humans outside Africa descend mainly from a single expansion out ∼50,000–70,000 years ago, but many details of this expansion remain unclear, including the history of the male-specific Y chromosome at this time.”
“A recent bottleneck of Y chromosome diversity coincides with a global change in culture” (2015) [Link]: “The estimated time line of the Y chromosome coalescent events in non-African populations (Supplemental Fig. S9) fits well with archaeological evidence for the dates of colonization of Eurasia and Australia by anatomically modern humans as a single wave ∼50 kya.”
CT's formation occurred 88,000 YBP [Link] and therefore preceded even the earliest estimated date of the migration (70,000 YBP), which means that it likely formed in Africa.
However, the haplogroups that descend from CT formed between 68,500 YBP and 65,200 YBP, which means that they could have formed either inside or outside of Africa, depending on the actual date that the migration occurred.
The argument that favors a Eurasian origin of E-M96 is that the AMHs who possessed CT supposedly migrated out of Africa and that the haplogroups that descend from CT therefore formed in their descendants outside of Africa; in accord with this argument, the presence of E-M96 inside of Africa is supposedly due to a back-migration. However, this is unlikely due to the most recent common ancestor (TMRCA) of E-M96 living further in the past relative to those of C-M130, F-M89, and D-CTS3946, which indicates that the carriers of the latter three haplogroups experienced at least one bottleneck event that the carriers of E-M96 did not experience; a bottleneck event is a natural disaster, disease outbreak, or anything else that eliminates a significant portion of a population - thereby restricting the common ancestor of that population to an individual who lived closer to the date of the bottleneck event. This is significant, because the migration out of Africa is associated with a bottleneck event [Link]; if E-M96 didn’t experience the bottleneck event that was associated with the migration out of Africa, then E-M96 was not carried out of Africa when C-M130, F-M89, and D-CTS3946 were carried out.
As can be seen below [Link], the most recent common ancestors of C-M130, F-M89, and D-CTS3946 lived only zero to 2,300 years apart. However, that of E-M96 lived 3,500 years to 5,800 years earlier.
C-M130: TMRCA 48,800 YBP
F-M89: TMRCA 48,800 YBP
D-CTS3946: TMRCA 46,500 YBP
E-M96: TMRCA 52,300 YBP
48,800 YBP - 48,800 YBP = 0 years
48,800 YBP - 46,500 YBP = 2,300 years
52,300 YBP - 48,800 YBP = 3,500 years
52,300 YBP - 46,500 YBP = 5,800 years
Additionally, the dates themselves imply that the bottleneck event occurred sometime between 46,500 YBP and 52,300 YBP, which overlaps with the most recent estimated date for the migration out of Africa: 50,000 YBP.
This is noteworthy, because the most recent common ancestor of E-M96 is the only one that lived before this date (52,300 YBP vs 50,000 YBP); this means that he would have lived before the migration and therefore would have lived inside of Africa - but the most recent common ancestors of C-M130, F-M89, and D-CTS3946 would have lived after the migration and therefore would have lived outside of Africa.
The logical conclusion to draw is that CT and the haplogroups that descend from it - C-M130, F-M89, D-CTS3946, and E-M96 - all formed inside of Africa, but that CT and / or the haplogroups that descend from it - with the exception of E-M96 - migrated out of Africa 50,000 YBP.
It should be noted that D0’s [D-Zero’s] presence in Africa is likely due to a remnant of males who carried D0 [D-Zero] or a precursor of it remaining in Africa, when the majority of males who carried subclades of D migrated out of the Africa 50, 000 YBP. The later date for TMRCA of D relative to the dates for TMRCAs of C and F may be due to the carriers of D who left Africa experiencing an additional bottleneck event as they migrated to East Asia, which is where D is carried at its highest frequencies.
Additionally, because there is no indication that C-M130, F-M89, or D-CTS3946 (with the exception of D0 [D-Zero]) ever spread throughout Africa, it is logical to conclude that they migrated out of Africa from the region where they must have formed: East Africa, because that is where the fossil record indicates that AMHs formed.
This means that E-M96 formed in East Africa as well, since it formed at the same time as D-CTS3946 - its “brother” haplogroup - and near the time at which C-M130 and F-M89 formed; however, rather than migrate out of Africa, it migrated further into the continent. Therefore, East Africa is the default location to consider as the birthplace of the Iberomaurusians’ original paternal haplogroup, E-M35, since it’s downstream relative to E-M96.
In regard to the nature of the population that spawned E-M35, it can be determined by considering the sub-Saharan African populations that share ancestry with the Taforalt specimens.
The Shared Source & The Direct Source
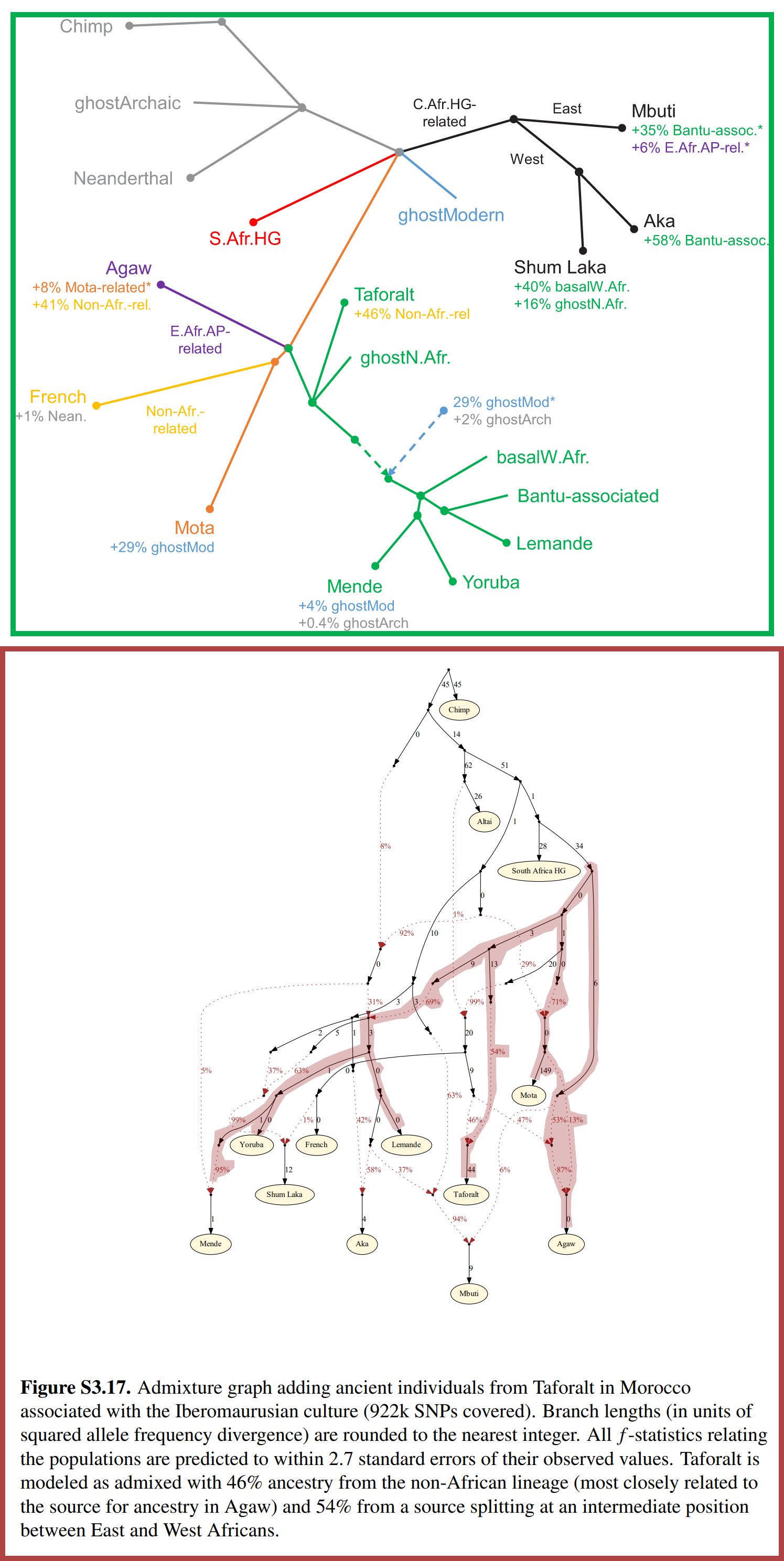
Figure 5 and Figure S3.17 of the peer-reviewed scientific article “Ancient West African foragers in the context of African population history” (2020) [Link] are admixture graphs that model the Taforalt specimens, other human fossils, and several modern populations. Note that the latter figure is included in the supplementary materials and not the main article [Link].
The relationships depicted in these admixture graphs are based on DNA analyses of human fossils, such the Taforalt specimens and the Ethiopian fossil Mota [Link], as well as several modern populations. They both depict the Taforalt specimens (labeled “Taforalt”) as deriving ancestry from the same source as Mota, the Agaw of Ethiopia, West Africans (namely the Mende, the Yoruba, and the Lemande), and Bantus.
This is noteworthy, because the Taforalt specimens*, Mota and the aforementioned modern populations are fully or predominantly sub-Saharan African - and they all possess paternal haplogroups that descend from E-M96. These details strongly indicate that the source of ancestry that is shared by them was also sub-Saharan African - and it carried a paternal haplogroup that descended from E-M96 and that was ancestral to their paternal haplogroups.
*Recall the SSA component of the re-expressed admixture model of the Taforalt specimens: 54.96% SSA + 45.04% Natufian
The value of 54.96% means that the Taforalt specimens are predominantly sub-Saharan African.
Note that the source of ancestry that is shared by The Taforalt specimens, Mota, the Agaw, West Africans and Bantus will be referred to as The Shared Source from hereon until it is identified.
Explanations About The Ancestries & Paternal Haplogroups Of The Taforalt Specimens, Mota, & Their Related Modern Populations
Below are explanations of the ancestries of the Taforalt specimens, the aforementioned fossil Mota, and the aforementioned modern populations - as well as their corresponding paternal haplogroups. The explanations make it clear that they are all fully or predominantly sub-Saharan African and possess paternal haplogroups that descend from E-M96, which indicate that The Shared Source was both sub-Saharan African and the source of their paternal haplogroups.
The Taforalt specimens
In the previous section, the Taforalt specimens were modelled as being 36.5% sub-Saharan African in the original two-way admixture model: 36.5% SSA + 63.5% Natufian
However, the admixture model was re-expressed after determining the amount of SSA ancestry in its Natufian component: 54.96% SSA + 45.04% Natufian
Note that the SSA component of the re-expressed admixture model is roughly equal to the amount of ancestry that the Taforalt specimens are modelled as deriving from The Shared Source in Figure S3.17: 54.96% ~= 54%. The difference of 0.96% is likely a margin of error that is the result of Figure S3.17 rounding values to whole intergers.
Therefore, The SSA component of the re-expressed admixture model is the ancestry that Figure S3.17 depicts The Taforalt specimens as deriving from The Shared Source, which means that The Shared Source was SSA (i.e. sub-Saharan African).
As already explained, the Taforalt specimens carry E-M78 (E1b1b1a1); additionally, it has been determined herein that the original paternal haplogroup of the Iberomaurusians - the population of the Taforalt specimens - was E-M35, which is ancestral to E-M78.
E-M96 -> E-M5479 -> E-P147 -> E-P177 -> E-P2 (E1b1) -> E-M215 (E1b1b) -> E-M35 -> E-L539 -> E-M78 (E1b1b1a1) [Link]
Given that the Taforalt specimens derive 54% to 54.96% of their ancestry from The Shared Source, it is unlikely that the Taforalt specimens don’t derive either their maternal or paternal haplogroup(s) from them. Therefore, given that they derive their maternal haplogroups form a Eurasian source, they likely derive their paternal haplogroup from The Shared Source.
Mota
Mota possesses no DNA from the migration of West Eurasians into East Africa 3000 YBP, which is explained in a quote that is from a peer-reviewed scientific article and that is further below. This indicates that Mota has no Eurasian DNA, since no previous migration of Eurasians into East Africa has ever been deduced or proven to have occurred. This implies that The Shared Source also did not possess any Eurasian DNA, since 71% of Mota's ancestry derived from The Shared Source. Therefore, both Mota and The Shared Source must have been fully sub-Saharan African.
Quote from “Ancient Ethiopian genome reveals extensive Eurasian admixture in Eastern Africa” (2015) [Link]: “By sampling the petrous bone, we sequenced the genome of a male from Mota Cave (herein referred to as “Mota”) in the southern Ethiopian highlands,… Mota’s remains were dated to ~4500 years ago [direct calibrated radiocarbon date] and thus predate both the Bantu expansion and, more importantly, the 3000-year-old West Eurasian backflow, which has left strong genetic signatures in the whole of Eastern and, to a lesser extent, Southern Africa…. As expected, we failed to find any West Eurasian component in Mota, thus providing support for previous dating of that event.”
Mota possesses Paternal Haplogroup E-Y175024.
E-M96 -> E-M5479 -> E-P147 -> E-P177 -> E-P2 (E1b1) -> E-V38 (E1b1a) -> E-M329 -> E-V2403 -> E-Y175024 [Link] (Mota’s ID is I5950)
Due to Mota being fully sub-Saharan African, all of his sources of ancestry - including The Shared Source - must have been sub-Saharan African.
Additionally, the 31% of Mota’s ancestry that does not come from The Share Source comes from an unknown population of modern humans (i.e. Homo sapiens), who are referred to as Ghost Modern in the aforementioned admixture graph Figure 5; they split into their own lineage 250,000 to 200,000 years ago, which is explained in a quote that is from a peer-reviewed scientific article and that is further below. Therefore, this population split long before the formation of E-M96 (65,200 YBP), which makes it unlikely to have been the source of Mota’s paternal haplogroup; this means that Mota must derive his paternal haplogroup from The Shared Source.
Quote from “Ancient West African foragers in the context of African population history” (2020) [Link]: “Recent consensus has been that southern African hunter-gatherers, who split from other populations ∼250,000–200,000 BP, represent the deepest sampled branch of modern human variation. Our results suggest that Central African hunter-gatherers split at close to the same time (perhaps slightly earlier)…. The modern human component diverges at almost the same point as Central and southern African hunter-gatherers and is tentatively related to the deep source contributing ancestry to Mota,…”
The Agaw
In the first admixture graph, the Agaw are labeled as inheriting 41% of their ancestry from non-Africans; this implies that they inherit the other 59% of their ancestry from the opposite of non-Africans (i.e. sub-Saharan Africans). This 59% SSA ancestry is modelled as coming from The Shared Source in the second admixture graph, which implies that The Shared Source is sub-Saharan African.
The 59% of their ancestry that’s from The Shared Source is not explicitly shown, because it’s combined with ancestry from other sources; therefore, it has to be calculated as follows:
87% of 53% -> 0.87(53%) = 46.11% -> 46.11% + 13% = 59.11% -> ~59%
The Agaw tested positive for E-P2 (E1b1) but tested negative for E-V38 (E1b1a), which means that they carry unidentified subclades of E-P2 (E1b1); the only other branch of E-P2 is E-M215 (E1b1b), which means that the Agaw carry subclades of E-M215. These results have been recorded in the doctoral thesis “Variation in Y chromosome, mitochondrial DNA and labels of identity on Ethiopia” (2011) [Link]. Their paternal haplogroup results are recorded on line 380 of columns Q & R of the thesis’ excel spreadsheet, in which they are listed as “AG” (an abbreviation of “Agaw”); 74 Agaw males who speak a Cushitic language and 52 Agaw males who speak a Semitic language tested positive for E-P2.
E-M96 -> E-M5479 -> E-P147 -> E-P177 -> E-P2 (E1b1) -> E-M215 (E1b1b) [Link]
The Agaw inherited the 41% of their ancestry that’s non-African from Arabians, which is why the Agaw also carry subclades of Paternal Haplogroup J [Link]; this is the signature paternal haplogroup of Arabians [Link]. The Agaws’ data for Paternal Haplogroup J is contained on line 479 of columns Q & R of the aforementioned thesis’ excel spreadsheet; 29 Agaw males who speak a Cushitic language and 23 Agaw males who speak a Semitic language tested positive for J.
The Agaw did not inherit Paternal Haplogroup E-M215 from their Arabian ancestors, since E-M215 is not a native haplogroup of Arabians; therefore, the Agaw must have inherited E-M215 from The Shared Source.
West Africans & Bantus
In the second admixture graph, West Africans are modelled as inheriting 69% of their ancestry from The Shared Source. Additionally, they possess no DNA from the migration of West Eurasians into East Africa 3000 YBP, and they have never been deduced or proven to possess Eurasian ancestry. Therefore, they are fully sub-Saharan African, which implies that The Shared Source was fully sub-Saharan African.
Quote from “Ancient Ethiopian genome reveals extensive Eurasian admixture in Eastern Africa” (2015) [Link]: “As expected, we failed to find any West Eurasian component in Mota, thus providing support for previous dating of that event.”
Quote from “Erratum for the Report ‘Ancient Ethiopian genome reveals extensive Eurasian admixture in Eastern Africa …” (2016) [Link]: “The Western Eurasian backflow mostly affected East Africa and only a few Sub-Saharan populations; the Yoruba and Mbuti do not show higher levels of Western Eurasian ancestry compared to Mota.”
The Yoruba are a West African ethnic group that are used in scientific studies to represent West Africans and Bantus.
The Mbuti are Central-African hunter-gatherers; however, they are not relevant to what is being presented herein.
West Africans and Bantus carry paternal haplogroups that descend from E-M132 [Link] and E-M2 [Link].
As with Mota, 31% of West Africans and Bantus' ancestry comes from the Ghost Modern population, who split into their own lineage 250,000 to 200,000 years ago; this is explained in another quote from the aforementioned peer-reviewed scientific article that is further below. Therefore, this population split long before the formation of E-M96 (65,200 YBP), which makes it unlikely to have been the source of West Africans and Bantus' subclades of E-M96; this means that said subclades must derive from The Shared Source.
Quote from “Ancient West African foragers in the context of African population history” (2020) [Link]: “The West African clade is distinguished by admixture from a deep source that can be modelled as a combination of modern human and archaic ancestry. The modern human component diverges at almost the same point as Central and southern African hunter-gatherers and is tentatively related to the deep source that contributes ancestry to the Mota individual,…”
The Ghost Population of Modern Humans
Mota's paternal haplogroup (E-Y175024) is a subclade of E-M329, which is the brother of West Africans and Bantus’ macro paternal haplogroup E-M2; both E-M329 and E-M2 are direct subclades of E-V38 (E1b1a). According to YTree's YFull, E-M329 and E-M2 both formed 39,200 years ago, which means that the lineage of Mota and that of West Africans and Bantus split from each other at that time. Therefore, because Mota, West Africans and Bantus all derive ancestry from the Ghost Modern population - and at the same percentage of 29% - their ancestors must have intermixed with the Ghost Modern population before they split into separate lineages 39,200 years ago.
Subsequently, because the lineages that produced E-M329 and E-M2 likely split from each other in East Africa, The Ghost Modern population must have been present in East Africa.
Quote from the peer-reviewed scientific article “A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms” (2011) [Link]: “Using the principle of the phylogeographic parsimony … in favor of a common ancestor of E-M2 and E-M329 strongly supports the hypothesis that haplogroup E1b1 originated in eastern Africa, as previously suggested, and that chromosomes E-M2, so frequently observed in sub-Saharan Africa, trace their descent to a common ancestor present in eastern Africa.”
Additionally, the aforementioned peer-reviewed scientific article, “Ancient West African foragers in the context of African population history” (2020) [Link], models the Ghost Modern population’s range of habitation as spanning from East Africa to West-Central Africa, along the southern edge of the Sahara; the model can be seen in Figure 4 of the article, which is posted below. Therefore, the Ghost Modern population will be referred to as Ancestral Saharans.

Figure 4 of the peer-reviewed scientific article, “Ancient West African foragers in the context of African population history” (2020) [Link].
Conclusions About The Shared Source & The Direct Source
Due to the Taforalt specimens, Mota, the Agaw, West Africans and Bantus all deriving ancestry from The Shared Source, and being predominantly or fully sub-Saharan African, and possessing paternal haplogroups that are subclades of E-M96 - conclusions that are being drawn herein are as follows:
The Shared Source was sub-Saharan African.
The Shared Source’s paternal haplogroup was also a subclade of E-M96.
The Iberomaurusians’ original paternal haplogroup, E-M35, descends from The Shared Source's paternal haplogroup; therefore, so do those of the Taforalt specimens and the Natufians, which were E-M78 and E-Z830, respectively.
The Natufians carried E-Z830, which is a paternal haplogroup that derived from The Shared Source - which has been determined herein to have been SSA; however, the Natufians’ amount of SSA ancestry decreased relative to the amount that their Iberomaurusian ancestors possessed (from ~54.96% to ~18.46%). This implies that the Natufians’ male lineage was comprised predominantly or entirely of males who possessed E-Z830 (or its precursor E-Z827) but that their female lineage was comprised predominantly or entirely of Eurasian females. This should not be surprising considering that the Natufians inhabited the Levant, which was predominantly inhabited by Eurasians, such as the proto-Arabians with whom they bred.
However, as a result of exchanging females with the proto-Arabians, the Natufians’ DNA - which included SSA DNA - introgressed into the gene pool of the proto-Arabians, and it did so maternally; this means that modern Arabians have Natufian ancestry - which includes pre-historic SSA ancestry - but they don’t have the Natufians’ paternal haplogroup (E-Z830) or any paternal haplogroup that descends from it (excluding any rare cases). This will be addressed later herein.
Due to E-P2 (E1b1) being the most downstream haplogroup relative to E-M96 that’s ancestral to the haplogroups of the Taforalt specimens, Mota and the aforementioned modern populations, a conclusion that is being drawn herein is that E-P2 (E1b1) was the The Shared Source’s paternal haplogroup.
However, because the Iberomaurusians’ original paternal haplogroup was E-M35 rather than E-P2 (E1b1), The Shared Source was not The Direct Source of the Iberomaurusians’ original paternal haplogroup; instead, The Shared Source was a precursor to The Direct Source.

Figure 5 and Figure S3.17 of the peer-reviewed scientific article “Ancient West African foragers in the context of African population history” (2020) [Link] - but with added labels that reflect the conclusions that are drawn below.
The origin of The Shared Source’s paternal haplogroup, E-P2 (E1b1), and the origin of the Iberomaurusians’ original paternal haplogroup, E-M35, were geographically linked, as indicated in the peer-reviewed scientific article “Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area” (2004) [Link]:
“Both phylogeography and microsatellite variance suggest that E-P2 and its derivative, E-M35, probably originated in eastern Africa. This inference is further supported by the presence of additional Hg E lineal diversification and by the highest frequency of E-P2* and E-M35* in the same region. The distribution of E-P2* appears limited to eastern African peoples. The E-M35* lineage shows its highest frequency (19.2%) in the Ethiopian Oromo but with a wider distribution range than E-P2*.”
Based on this quote, both the paternal haplogroups of The Shared Source and The Direct Source - which were E-P2 (E1b1) and E-M35 - originated in East Africa; to be specific, they originated in Ethiopia, which is in the Horn of Africa. Therefore, conclusions that are being drawn herein are the following:
The Shared Source inhabited the Horn of Africa.
Both paternal haplogroups E-P2 (E1b1) and E-M35 were carried by The Shared Source.
A subset of The Shared Source - whose males carried E-M35 - split off and migrated to North Africa. This must have happened, in order for carriers of E-M35 to intermix with the Aterian natives of North Africa and the Eurasian migrants therein - and thereby create the Iberomaurusians, who were also native to North Africa.
It cannot be determined whether Eurasian migrants intermixed with the Aterian natives of North Africa before E-M35 carriers arrived therein - or whether E-M35 carriers intermixed with the Aterian natives of North Africa before Eurasian migrants arrived therein. However, The Direct Source was the first combination of these populations that included E-M35 carriers.
Assuming that Paternal Haplogroups E-V38 (E1b1a) and E-M215 (E1b1b) also formed in the Horn of Africa, they would have also been carried by The Shared Source.
Therefore, The Shared Source will be referred to as AHAs (Ancestral Horn Africans) and The Direct Source will be referred to as ANAs (Ancestral North Africans) - the latter of which was coined by Iosif Lazaridis [Link] in a scientific article that's mentioned later herein.
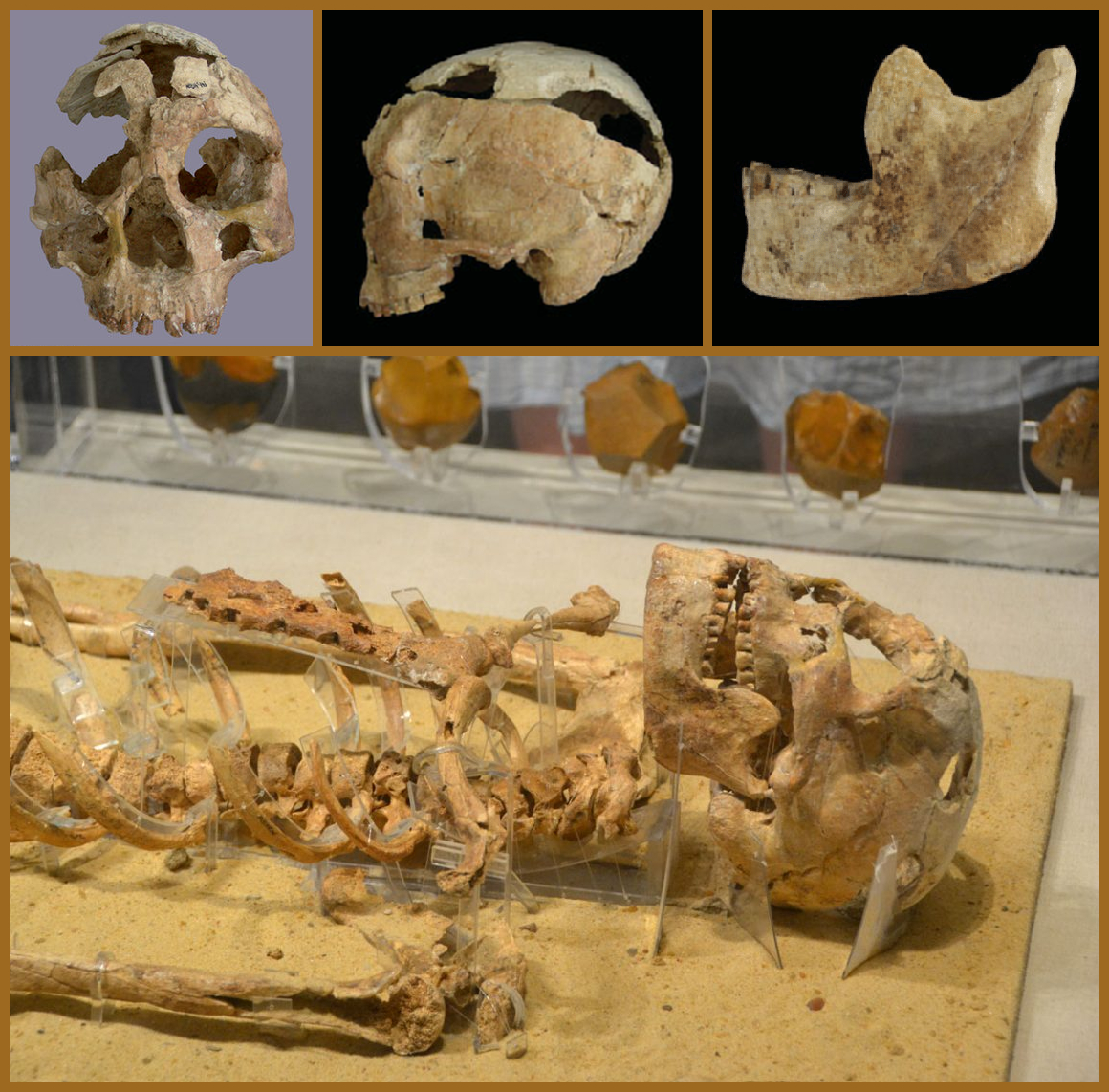
These are photos of Nazlet Khater 2 [Link], which is the fossilized skeleton of a man who lived in Egypt 35,000 years ago.
Given the location and age of this individual, he was likely an Ancestral North African.
He was discovered in Egypt, which is in North Africa.
He has been estimated to have lived 33,000 years ago, which was 1,700 years after the formation of the Iberomaurusians’ first paternal haplogroup: E-M35 [Link]. Therefore, his paternal haplogroup may have been E-M35.
His mandible has very wide rami and a tall main body, which are characteristics of Aterians and Iberomaurusians, as explained earlier herein; however, his morphology is otherwise anatomically modern. Therefore, he was likely an Aterian-AMH hybrid.
Given that it has been concluded herein that the Iberomaurusians were Aterian-AMH hybrids, it is logical that an Ancestral North African would have been such a hybrid as well - because the Ancestral North Africans were ancestral to the Iberomaurusians.

A human ancestry graph that provides a graphic representation of conclusions that have been drawn herein and of the aforementioned admixture graphs, Figure 5 and Figure S3.17.

Figure 2. from “Paleolithic DNA from the Caucasus reveals core of West Eurasian ancestry” (2018) [Link] is an admixture graph that models Paleolithic West Eurasians. As can be seen, it includes Ancestral North Africans as one of the populations from which the Taforalt specimens derive ancestry.
As does the aforementioned Figure S3.17 [Link], this admixture graph models the Taforalt specimens, Mota, and West Africans (and Bantus) - the last of whom are represented by the Yoruba - as deriving ancestry from a source that they share with one another. However, it models the Taforalt specimens and West Africans (and Bantus) as deriving only 45% and 13% of their ancestries, respectively, from this source - whereas the aforementioned Figure S3.17 models them as deriving 54% and 69% of their ancestries, respectively, from it.
Additionally, it models West Africans (and Bantus) as deriving their ancestry from this source via the lineage of the Taforalt specimens, rather than independently.
Given that the ancestors of West Africans and Bantus did inhabit the Sahara when it was comprised of grasslands, forests, and lakes 8000 years ago and prior [Link], it is likely that they did intermix with Taforalt-related populations (e.g. the Kiffians [Link] and Tenerians [Link]), because such populations also inhabited the Sahara at that time.
However, this possibility does not contradict the aforementioned admixture graphs, Figure 5 and Figure S3.17. West Africans and Bantus’ macro paternal haplogroup E-M2 is proof that they also independently descend from a source that contributed ancestry to the Taforalt specimens, for the following reasons:
E-M2 and the Taforalt specimens’ E-M78 both descend from a common ancestral haplogroup (E-P2 / E1b1), and the lineages of E-M2 and E-M78 split from one another approximately 41,400 years ago, when their ancestors - E1b1a and E1b1b, respectively - formed, according to YFull’s YTree.
That’s 16,400 years before the Taforalt specimens’ Iberomaurusian culture appeared approximately 25,000 YBP, which means that the lineage that stems from this source and that produced West Africans and Bantus’ split from the lineage that produced the Iberomaurusians long before the Iberomaurusians even existed as a distinct people.
E-P2 (E1b1) -> E-V38 (E1b1a) ->E-M2 [Link]
E-P2 (E1b1) -> E-M215 (E1b1b) -> E-M35 -> E-L539 -> E-M78 (E1b1b1a1) [Link]
Additionally, that West Africans and Bantus derive ancestry from this source via the lineage of the Taforalt specimens supports the hypothesis that West Africans and Bantus derive ancestry from the Aterians [Link], since it has been concluded herein that the Iberomaurusians descend from the Aterians.

Figure 2 of the peer-reviewed scientific article, “Pleistocene North African Genomes Link Near Eastern And Sub-Saharan African Human Populations” (2018) [Link].
In accord with the first two admixture graphs that have been shown herein [Link], this principle component chart shows that the Taforalt specimens are positioned intermediately relative to West Africans and Eurasians.
This is consistent with a quote from the article that features this chart.
“West Africans, such as Mende and Yoruba, most strongly pull out the sub-Saharan African ancestry in Taforalt.”
However, despite the Taforalt specimens and West Africans being modelled in the second admixture graph [Link] as inheriting 54% and 69% of their ancestries from the same source (AHA), respectively - the Taforalt specimens cluster closer to Eurasians than they do to West Africans.
A possible explanation for the Taforalt specimens’ closer proximity to Eurasians relative to West Africans
AHAs were genetically closer to Eurasians than they were to the other ancestries in West Africans (which are labeled “Ghost Modern” and “Ghost Archaic” in the first admixture graph [Link]). However, AHAs were not the same as Eurasians; if they were so, then the 54% of the Taforalt specimens’ ancestry that is from AHAs - combined with the other 46% of their ancestry that’s from Eurasians - would push the Taforalt specimens into the genetic isolate formed by Eurasians.
AHAs were genetically closer to Eurasians, due to sharing ancestors with Eurasians that they did not share with the other populations from whom West Africans descend (“Ghost Modern” and “Ghost Archaic”). This is evident due to AHA’s paternal haplogroups (e.g. E-P2 [E1b1], E-M35, etc) being downstream relative to Paternal Haplogroup CT, as are the paternal haplogroups of Eurasians.
AHAs - and all sub-Saharan Africans whose paternal haplogroups are downstream relative to E-M96 - descend from the proto-Out-of-Africa population. However, when the males who carried paternal haplogroups that were downstream relative to C-M130, F-M89, and D-CTS3946 migrated out of Africa 50,000 YBP, those who carried paternal haplogroups that were downstream relative to E-M96 stayed in Africa.
Many sub-Saharan African populations that carry paternal haplogroups that are downstream relative to E-M96 also carry maternal haplogroups that are downstream relative to L3, which spawned the maternal haplogroups of Eurasians. This strongly indicates that these populations descend from the proto-Out-of-Africa population and are therefore genetically closer to Eurasians than other sub-Saharan Africans.
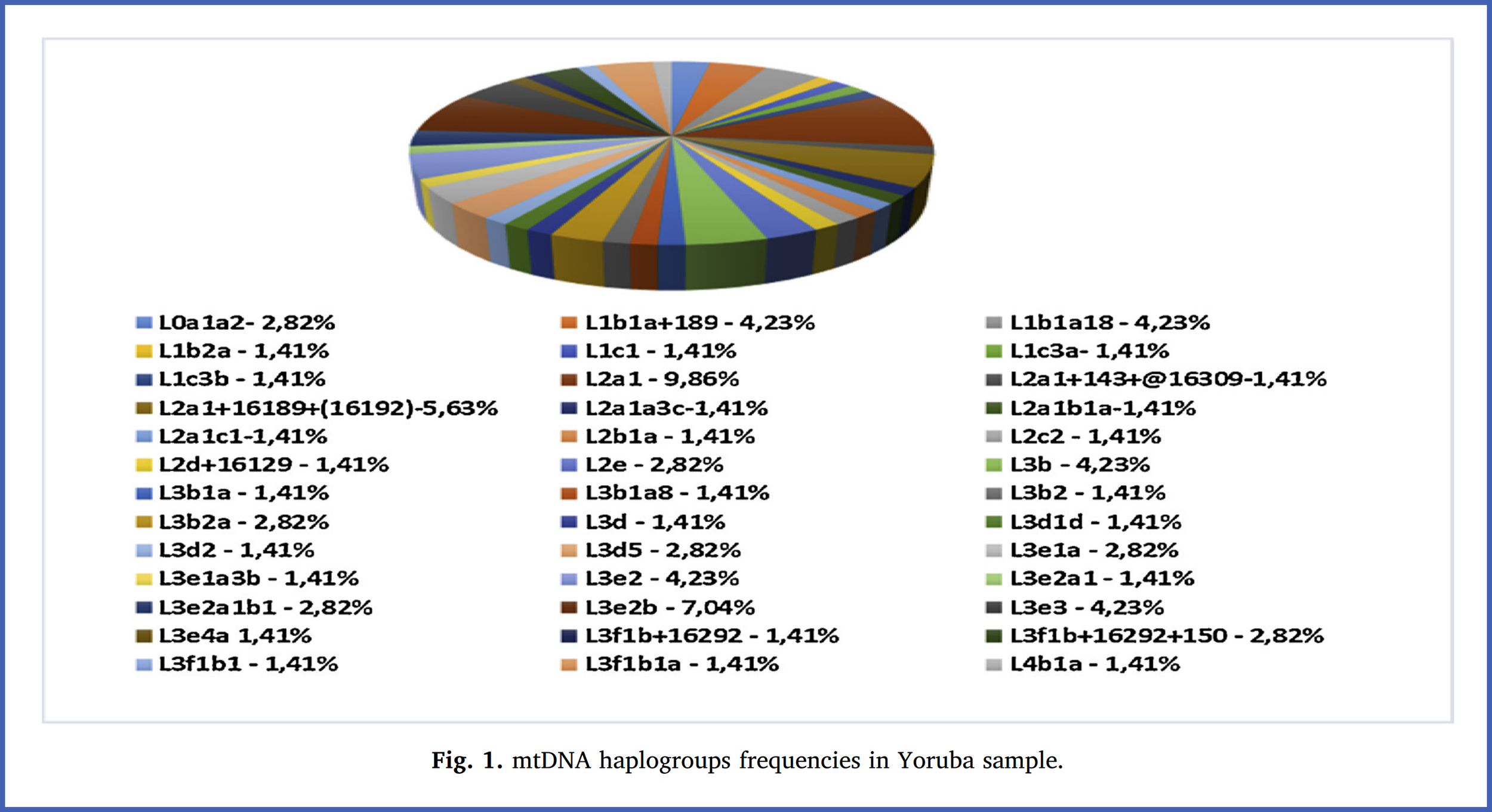
Figure 1 of the peer-reviewed scientific article “Mitochondrial genetic profile of the Yoruba population from Nigeria” (2019) [Link].
The figure shows that subclades of Maternal Haplogroup L3 - which spawned the maternal haplogroups of Eurasians - are quite abundant among the Yoruba of Nigeria, who are West Africans. This is noteworthy, because the paternal haplogroups of the Yoruba are downstream relative to E-M96 - which means that both the maternal and paternal haplogroups of the Yoruba indicate that they descend from the proto-Out-of-Africa population (but their ancestors split from it and remained in Africa); therefore, the Yoruba would be among the sub-Saharan Africans who are the most closely related to Eurasians (minus admixture that other sub-Saharan Africans may have from the latter, which would inflate their relatedness). Other such sub-Saharan Africans include Mota and the aforementioned populations with whom the Taforalt specimens share ancestry: the Agaw, [other] West Africans and Bantus - as well as the Oromo.
Mota’s maternal haplogroup is L3x2a2b* [Link] (Mota’s ID is I5950)
West Africans, such as the Yoruba, possess various subclades of L3 as can be seen in the figure above.
Bantus possess subclades of L3, including but not limited to L3e1 and L3e4.
Quote from “Phylogenetic and phylogeographic analysis of African mitochondrial DNA variation” (2004) [Link]: “Further evidence for the influence of the Bantu movements on the genetic landscape of Africa is widely reported; the broad distributions of the haplogroups L3e1 and L3e4, for example, are reported to be the due to the Bantu dispersals (Bandelt et al 2001).”
The Oromo posses subclades of L3, such as L3x.
Quote from the peer-reviewed scientific article “Ethiopian Mitochondrial DNA Heritage: Tracking Gene Flow Across and Around the Gate of Tears” (2004) [Link]: “Haplogroup L3x is defined by transitions at nps 6401, 13708, and 16169 and was found in 10 Ethiopians, with most frequent appearance (12%) among the Oromos.”
Another possible explanation for the Taforalt specimens’ closer proximity to Eurasians relative to West Africans
The As already explained, the Natufians’ SSA ancestry introgressed into the genepool of the proto-Arabians. Therefore, there is pre-historic SSA ancestry in modern Arabians, who are a subset of Eurasians; this causes the distance between Eurasians and the Taforalt specimens - from whose population the Natufians inherited this SSA ancestry - to be less than it would be otherwise. This is reflected in the original two-way admixture model of the Taforalt specimens, which explicitly defines 36.5% of their ancestry as SSA - but hides the remaining 18.46% SSA ancestry in the 63.5% Natufian component, as already explained; this 18.46% SSA ancestry is the portion that introgressed into the genepool of the proto-Arabians and is present in the genepool of modern Arabians. Therefore, 45.04% of the Taforalt specimens’ ancestry pulls towards Eurasians (63.5% - 18.46% = 45.04%), while only 36.5% pulls towards West Africans.
The hidden 18.46% SSA ancestry pulls towards both West Africans and Eurasians - thereby being effectively neutral - because it's present in both sets of populations.
The following two images prove that Arabians have the most Natufian ancestry and therefore the most affinity to sub-Saharan Africans (due to the SSA ancestry that Arabians have inherited through their Natufian ancestry). It is logical to conclude that this affinity is what causes The Taforalt specimens to cluster closer to Eurasians than to West Africans; the SSA ancestry in Arabians partially negates the pull of the SSA ancestry in the Taforalt specimens toward West Africans.

Here are the genetic distances between Arabians and sub-Saharan Africans - and those between other Middle Easterners and sub-Saharan Africans. They were calculated using the Vahaduo Admixture Calculator [Link] and G25 DNA Coordinates from Eurogenes [Link].
Arabians (inhabitants of the Arabian Peninsula): Saudi, SaudiA, SaudiB, BedouinA, BedouinB, and Yemenites.
Other Middle Easterners: Iranian, Iraqi, Jordanian, Palestinian, and Syrian.
Sub-Saharan Africans: West Africans: Mende Sierra Leone, Yoruba, Esan Nigeria; Central Africans: Biaka and Mbuti; and East Africans: Ethiopian Ari and Hadza; Southern Africans: Khomani San.
Note that the smallest distances between Arabians and sub-Saharan Africans are approximately half of the smallest distances between other Middle Easterners and sub-Saharan Africans, which is consistent with Arabians having more Natufian ancestry - and therefore more SSA ancestry. This is noteworthy, because Arabians have the most Natufian ancestry out of all Middle Easterners, as can be seen in the next image.

Note that the Arabian populations have the highest levels of Natufian ancestry (16% to 27.8%), but the other Middle Eastern populations have lower amounts of Natufian ancestry (0% to 6.2%). This correlates with the distances between these population and sub-Saharan Africans, which are shown in the previous image; the more Natufian ancestry that a population has, the less genetic distance there is between it and sub-Saharan Africans.
Furthermore, most Arabian males carry paternal haplogroups that are downstream relative to Paternal Haplogroup J (J-M304), rather than any that are downstream relative to E-M96. This relates to a hypothesis that was presented herein, which is that Natufian males consistently exchanged females with proto-Arabians and that therefore modern Arabians inherited their Natufian ancestry - which includes SSA ancestry - maternally; subsequently, modern Arabians have Natufian autosomal DNA but do not have Natufian or Natufian-derived paternal haplogroups (excluding rare exceptions).
Aterian Ancestry In the Taforalt Specimens

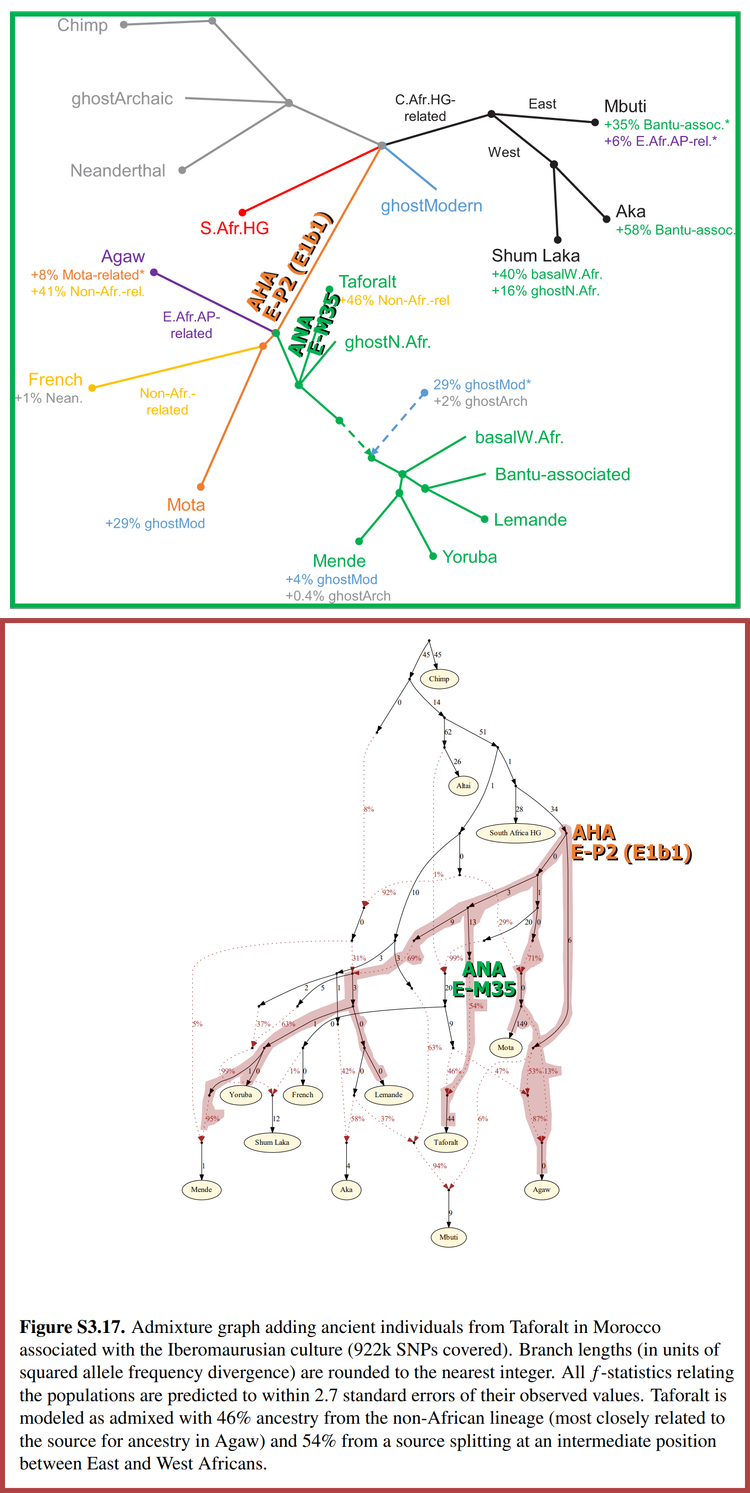
Figure S3.17 of the peer-reviewed scientific article “Ancient West African foragers in the context of African population history” (Supplementary Materials) (2020) [Link].
A conclusion that has been made herein is that the Taforalt specimens were hybrids of AMHs and Aterians (who have been concluded herein to have been another subspecies of Homo sapiens). Therefore, part of the 18.46% SSA ancestry of the Taforalt specimens that is hidden in the original two-way admixture model could be Aterian ancestry rather than SSA ancestry. This could be due to some software programs categorizing any DNA that is not Eurasian as sub-Saharan African, or it could be due to Aterian DNA having introgressed into the genepools of West Africans [Link] and therefore being interpreted as SSA ancestry by software programs.
Given that a portion of this 18.46% ancestry includes 6.8% Omotic ancestry, the Aterian ancestry could be the other, unspecified 11.66% SSA ancestry (18.46% - 6.8% = 11.66%).
This 11.66% ancestry wouldn’t have been inherited from AHA, which means that only 43.3% of the 54% to 54.96% ancestry that the Taforalt specimens are modelled as inheriting in Figure S3.17 would have actually come from AHA. However, this does not contradict Figure s3.17, because of how it’s structured.
Note that in Figure S3.17 there is a downward-pointing black arrow above the text that reads “ANA E-M35,” and besides it is the number 13 (a red circle has been drawn around them in order to highlight them). The arrow and number represent genetic drift or other events that genetically differentiate a population from a source of its ancestry; in this case, the arrow and number could represent - at least partially - Aterian DNA that introgressed into the lineage that produced ANA. Therefore, the original two-way admixture model can be re-expressed in another form:
36.5% SSA + 6.8% SSA [Omotic] + 45.04% Natufian + 11.66% Aterian = 43.3% SSA + 45.04% Natufian + 11.66% Aterian
If this DNA is indeed Aterian, then modern North Africans (who descend from the Iberomaurusians) - as well as modern Arabians and other populations into which the 18.46% ancestry has introgressed - are partially Aterian.

{kind=link}
{kind=link}
As can be seen, modern North Africans have retained very large percentages of ancestry from the Iberomaurusians (an average of 22.4%), which means that there has been population continuity in North Africa for at least 15,000 years. Additionally, if the Iberomaurusians descended from the Aterians and if the Aterians descended from the Jebel-Irhoud humans, then there has been population continuity in North Africa for the past 300,000 years! This is very, very impressive and is a testament to the ancestral ties that modern North Africans have to the region.