The Hominin Lineage: Deducing the Phylogenetic Position of Yunxian 2 & the Identity of the Last Common Ancestor
Organisms are divided into topological groups via the field of biological taxonomy, with higher positioning meaning greater inclusivity and lower positioning meaning less inclusivity; for example, a tribe is more inclusive than a genus, which - in turn - is more inclusive than a species. Typically, the formal designation of an organism is a binomial term that begins with their genus and ends with their species, genus being the directly higher group-classification relative to species. An example of this is the formal designation of Modern Humans: Homo sapiens. Homo specifies our genus and sapiens specifies our species. This raises a question: What other species of Homo are there? Unfortunately, there are none. However, this has not always been so, as has been revealed by countless discoveries of fossilized Homo remains.
Such discoveries have revealed the past existences of Homo ergaster, Homo erectus, Homo heidelbergensis, Homo longi, Homo neanderthalensis and a few other species whose status as Homo are contested (e.g. Homo habilis, Homo floresiensis).

A topological tree that depicts the lineage of the genus Homo. Kingdom, Phylum, Class, etcetera are topological classifications; and the terms that precede them (e.g. Animalia, Chordata, Mammalia, etc) are the specific topological groups that form the ancestral lineage of Homo.
Homo belongs to the subtribe Australopithecina (aka Hominina), which split from the subtribe of Chimpanzees and Bonobos (i.e. Panina) ~7 million years ago; the lineage of gorillas split a few million years earlier, and that of orangutans split even earlier. Australopithecina includes the genera (plural of genus) Australopithecus and Paranthropus - the former of which was ancestral to Homo and the latter of which was a sibling lineage. Species of Australopithecus were similar to panins (members of Panina) but were adapted to both arborealism and bipedalism, and they had slightly larger brains than panins.
The traditional consensus regarding the phylogeny that produced Homo and subsequently modern humans is as follows:
Australopithecus → Homo habilis → Homo ergaster & Homo erectus | Australopithecus → Homo habilis → Homo ergaster → Homo erectus
Homo ergaster → Homo heidelbergensis
Homo heidelbergensis → Neandersovans
Neandersovans → Homo neanderthalensis & Asian Neandersovans
Asian Neandersovans + Asian Homo erectus → Homo longi / Denisovans
Homo heidelbergensis → Homo sapiens
However, recent analysis of an ancient hominin skull is said to challenge this phylogeny. The skull is named Yunxian 2 and was discovered in the Yunyang District of China’s Hubei Province; and the analysis on it was conducted by a team of researchers that was led by Professor Xiaobo Feng of Beijing Union University. The team’s methodologies and conclusions are presented in the peer-reviewed article, The phylogenetic position of the Yunxian cranium elucidates the origin of Homo longi and the Denisovans (2025) [peer-reviewed version: link] [non-peer-reviewed version: link]; the team will be referred to herein as Feng et al, because it was led by Professor Feng.

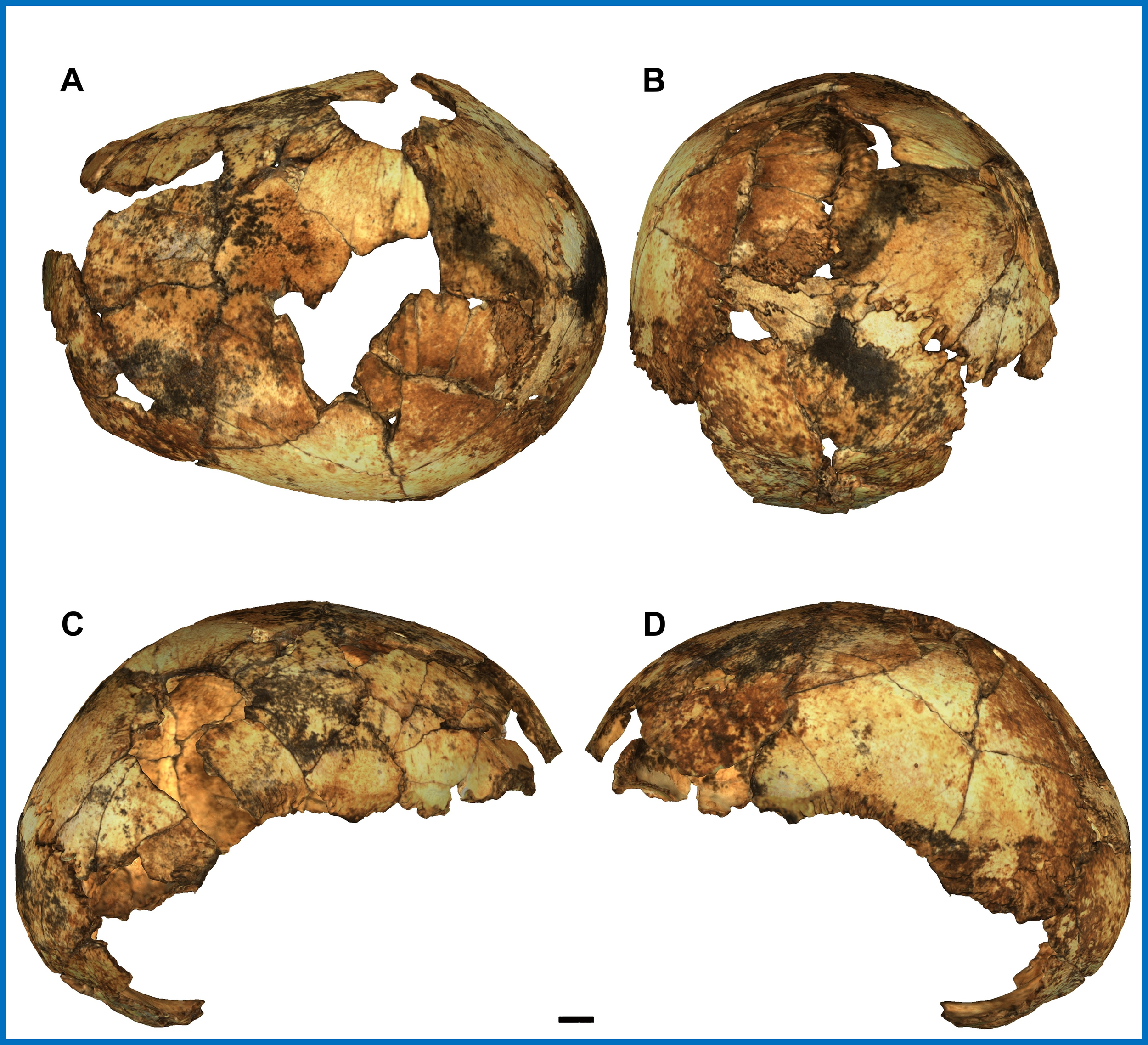
Figure 1: A composite of images showing the virtual reconstruction of Yunxian 2 from different angles
Feng et al’s methodologies essentially entailed comparing Yunxian 2’s physical traits and estimative age to those of other hominin skulls, and subsequently drawing conclusions about Yunxian 2’s position in the hominin phylogeny relative to those other skulls.
Among Feng et al’s conclusions is that Yunxian 2 is not a Homo erectus specimen, despite dating back to approximately 1 million years ago - when Homo erectus was the most widespread and advanced species of Homo according to traditional consensus. Some of Feng et al’s reasons for this conclusion are that - compared to Homo erectus - Yunxian 2’s brain case is smoother due to lacking sharp angles (particularly in the occipital region), its mastoid processes (the regions of the skull above the ears) are larger, its brow ridges are smaller and less pronounced, its midface projects forward to a lesser degree, the plane of its zygomaxillary region faces forward, and its third molars are smaller.
Note that Yunxian 2 and the other two crania with which it was discovered have been estimated to be 1.77 Ma in another scientific article: The oldest in situ Homo erectus crania in eastern Asia: The Yunxian site dates to ~1.77 Ma [Hu et al (2026)] [link]. However, this older age has not been unanimously accepted by the paleontology community, because the dating method that determined it is novel. Furthermore, Yunxian 2’s brain size is closer to that of younger African and Asian fossils (e.g. OH 9, the Daka cranium, Java Man, etc) rather than fossils that date back to 1.77 Ma ago (i.e. Dmanisi hominins and Homo ergaster specimens); said African and Asian fossils will be discussed in detail later herein.

Profiles of Yunxian 2 Reconstruction and Homo erectus (Peking Man) Reconstruction
Feng et al reached another conclusion by comparing very complex measurements of Yunxian 2, 24 ancient hominin skulls, and 153 recent human skulls; the conclusion was that Yunxian 2 clusters with several particular hominin skulls that were discovered in China, such as Dali Man, Jinniushan human, and the Harbin cranium. This is significant, because these skulls had already been determined to comprise a clade based on their strong morphological similarities, namely the Homo longi clade. Furthermore, protein analysis of one of these skulls, namely the Harbin cranium, revealed that it’s a Denisovan (source). Therefore, Homo longi and Denisovans may be the same species, or - at the very least - Denisovans may be a derivation of Homo longi. Given that Yunxian 2 has been estimated to be 1 million years old, Homo longi - including the lineage that produced Denisovans - would be at least the same age, or so Feng et al posits.
This challenges the traditional phylogeny of hominins, because said phylogeny posits that Denisovans formed as an independent lineage only 700,000 to 800,000 years ago, when they and Neanderthals split from a common lineage.
However, it must be considered that even though Homo longi is at least 1 million years old, the lineage that produced Denisovans may be younger. This is because genetic analyses indicate that Denisovans - and Neanderthals - derived some of their aDNA (autosomal DNA) from a superarchaic lineage; such a lineage is one that would have split over 1 million years ago from that which produced the LCA (Last Common Ancestor) of Denisovans, Neanderthals and Modern Humans, based on estimative divergence dates of certain Denisovan and Neanderthal gene variants. In other words, the Neanderthal and Denisovan lineages formed when a superarchaic lineage engaged in admixture events with the Neandersovan subgroup of the LCA; Neandersovans would have been the subgroup of the LCA that was ancestral to Denisovans and Neanderthals to the exclusion of Modern Humans.
Such admixture events are explained in the peer-reviewed article, Neanderthal-Denisovan ancestors interbred with a distantly related hominin (2020) [link]. The lead author of this article is Alan R. Rogers of University of Utah’s Department of Anthropology, so the authors of the article will be referred to herein as Rogers et al.
Roger et al’s analyses indicate that Neandersovans admixed with a superarchaic lineage, and their admixed descendants split into two lineages millennia later: ancestors of Neanderthals and ancestors of Denisovans. Finally, the ancestors of Denisovans further admixed with the superarchaic lineage, resulting in Denisovans carrying additional superarchaic admixture. This is represented in Figure 1 of Roger et al’s article, which is shown below.

Figure 1 of the article, Neanderthal-Denisovan ancestors interbred with a distantly related hominin (2020)
In the following excerpt from their article, Rogers et al proposes dates for the emergence of the superarchaic lineage:
Our results revise the date at which superarchaics separated from other humans. One previous estimate put this date between 0.9 and 1.4 Ma [(2), p. 47], which implied that superarchaics arrived well after the initial human dispersal into Eurasia around 1.9 Ma. This required a complex series of population movements between Africa and Eurasia [(33), pp. 66 to 71]. Our new estimates do not refute this reconstruction, but they do allow a simpler one, which involves only three expansions of humans from Africa into Eurasia: an expansion of early Homo at about 1.9 Ma [million years] ago, an expansion of neandersovans at about 700 ka ago, and an expansion of modern humans at about 50 ka ago.
This excerpt presents two scenarios:
Homo first arrived in Eurasia 1.9 Ma ago, and a second wave of Homo arrived in Eurasia between 0.9 and 1.4 Ma ago and produced the superarchaic lineage
Homo first arrived in Eurasia 1.9 Ma ago and produced the superarchaic lineage
Even though Rgers et al presents the second scenario as an alternative, they explicitly state that it’s just a simplification and that their results do not refute the first scenario. This is an important detail; it will factor into determining herein when the superarchaic lineage likely arrived in Eurasia.
Given that Homo longi formed at least 1 million years ago, it would have been part of the superarchaic lineage - stemming from either an expansion of Homo that occurred 0.9 to 1.4 Ma ago or as early as 1.9 Ma ago. Furthermore, the additional superarchaic admixture in Denisovans would explain the morphological similarity between Yunxian 2 and Denisovans. This raises the question: From which ancient African species or genus did the superarchaic lineage split?
Answering this question entails exploring Eurasian hominin fossils, the oldest of which are the Dmanisi hominins (link). These hominin fossils include 5 skulls and over 100 post-cranial remains that are approximately 1.7 million years old. Some of the Dmanisi fossils appear to be from australopithecines or Homo habilis (which is considered to be a late australopithecine species by some despite its classification), but others appear to be from the more derived species Homo erectus. This is explained in the peer-reviewed article, Testing the taxonomy of Dmanisi hominin fossils through dental crown area [aka Nery et al (2025)] [link].
… Considering the value of dental crown area in reconstructing evolutionary relationships, a comparative sample of fossil hominins, consisting of 51 maxillary and 71 mandibular specimens (583 teeth in total), was analyzed using Linear Discriminant Analysis (LDA) to evaluate the diversity in the Dmanisi fossil assemblage. Morphological affinities were examined visually through the first two discriminant functions, and taxonomic relationships were tested via classification analyses based on posterior probabilities. The analyses show a strong association of the D4500-D2600 (link) specimen with australopiths, and of the D2282-D211 (link) and D2700-D2735 (link) specimens with Homo species. … We conclude that differences in crown dimensions support the hypothesis of two distinct taxa coexistent at the Dmanisi site, previously proposed to be Homo georgicus and Homo caucasi. This proposal has important implications for the dispersal of Homo out of Africa at the beginning of Pleistocene. - Nery et al (2025)
Because skulls D2280 (link) and D3444-D3900 (link) lack teeth, they were not included in the analysis that’s explained above. However, their cranial morphologies and thick brow ridges are clearly those of Homo erectus. Therefore, there’s the possibility that the superarchaic lineage was an expansion of australopithecines, Homo habilis or Homo erectus from Africa into Eurasia.
The first possibility means that Homo longi would have evolved directly from australopithecines in Eurasia. However, this would have been unlikely, because it would have entailed two separate lineages of australopithecines improbably evolving into Homo in both Africa and Eurasia - given that the fossil record strongly indicates that australopithecines evolved into Homo ergaster in Africa.
The second possibility means that Homo longi would have evolved from Homo habilis in Eurasia. This seems more probable due to habilis being Homo; it’s classified as such because it exhibits a larger brain capacity, smaller jaws and weaker prognathism than australopithecines - which can be observed via the following list of several australopithecine and Homo habilis specimens.
Australopithecus afarensis AL 444-2 (link): age = 3 Ma | brain capacity: 550 cc
Australopithecus africanus Taung 1 (link): age = 2.8 Ma | brain capacity: 405 cc
Australopithecus africanus Sts 5 (link): age = 2.5 Ma | brain capacity: 485 cc
Australopithecus africanus Stw 505 (link): age = 2.5 Ma | brain capacity: 515 cc (Table 1 of Lockwood et al (1999): link)
Homo habilis KNM-ER 1813 (link): age = 1.9 Ma | brain capacity: 510 cc
Homo habilis OH 24 (link): age = 1.8 Ma | brain capacity: 600 cc
Homo habilis KNM-ER 1805 (link): age = 1.7 Ma | brain capacity: 600 cc (Text of Britannica article: link)
However, Homo habilis retained many post-cranial Australopithecine traits (e.g. proportionally long arms, short stature and overall small body size), so it’s unlikely that two geographically separated lineages of Homo habilis would have developed the even larger brain capacities and even weaker prognathism of Homo ergaster and Homo longi.
The probable scenario is that the superarchaic lineage was an expansion of Homo erectus, who were transcontinental and whose morphology is quite similar to that of the strictly African species Homo ergaster. The main differences between the two species are their degrees of robusticity; Homo ergaster specimens have thinner cranial bones, smaller or absent sagittal keels, smaller and less pronounced brow ridges, rounder crania, and a smaller range of brain capacities (i.e. 600 to 900 cc vs 800 to 1100 cc); however, the two species are otherwise similar. This strongly indicates that Homo ergaster and Homo erectus are very closely related, which is why they’re actually considered the same species by some anthropologists. Furthermore, that Homo erectus is essentially a more robust variation of Homo ergaster indicates that Homo ergaster may have evolved into Homo erectus; the larger range of brain capacities exhibited by Homo erectus supports this hypothesis, given that the brain capacity of Homo has generally increased over time.
The oldest Homo ergaster fossil is DNH 134, which is a calvaria (the top portion of the brain case) that was discovered in South Africa. It’s been determined to have belonged to a 2 or 3-year-old juvenile who lived 2 million years ago; this indicates that Homo ergaster existed early enough to have expanded into Eurasia between 0.9 and 1.9 Ma ago, which was when the superarchaic lineage expanded into Eurasia according to Rogers et al.
The DNH 134 Calvaria
Despite it being only part of a skull, it’s recognizably a Homo ergaster specimen. From a lateral view, its posterior curves downward at an angle that’s consistent with the oval shape of a Homo ergaster cranium; conversely, the crania of australopithecines and Homo habilis are rounder and broader at their posteriors from the same viewing angle. Additionally, the degree to which the top of the calvaria is narrower at its anterior than it is at its posterior (i.e. post-orbital constriction) is not as extreme relative to australopithecines and Homo habilis.
The second oldest and subsequent fossils of Homo ergaster are 1.75 to 1.6 million years old, and they include several skulls and ~40% of a skeleton (i.e. Turkana Boy).
Homo ergaster KNM-ER 3733 (link): age = 1.75 Ma | brain capacity: 850 cc
Homo ergaster KNM-ER 3883 (link): age = 1.6 Ma | brain capacity: 800 cc (Text of Garvin et al (2017))
Homo ergaster KNM-WT 15000 [Turkana Boy] (link): age = 1.6 Ma | brain capacity: 880 cc
Their ages make them slightly older or roughly contemporaneous with the aforementioned Dmanisi hominins. Recall that most of the Dmanisi hominins have been determined to be Homo erectus; so their age of ~1.7 Ma makes them the oldest specimens of their species.
The next oldest Homo erectus specimens are African:
Olduvai Hominid 9 (OH 9): age = 1.4 Ma | brain capacity = 1000 cc
BOU-VP-2/66 (Daka skull): age = 1 Ma | brain capacity = 1000 cc (Tattersall (2023))
Regarding the differences in the ages and brain capacities of the Dmanisi and African Homo erectus specimens, they seemingly indicate that a lineage of Homo ergaster migrated to Eurasia, evolved into Homo erectus therein and returned to Africa. However, morphological analyses indicate otherwise.
In particular, we find that Daka shares the greatest affinities with crania such as KNM-ER 3733 and 3883 from Koobi Fora, Turkana (dated to about 1.6 Ma), and it is farther from the far eastern Asia hypodigm. Moreover, two of the Dmanisi hominids, Georgia (1.7–1.8 Ma), the OH9 partial braincase from Olduvai (approximately 1.2 Ma), and an example of the late H. erectus calvaria from Ngandong (less than 100 ka) have somewhat intermediate phenotypes, consistent with the fact that, in different ways, these fossils are detached chronologically and/or geographically from typical H. erectus specimens from Java and China. Similarly, Daka is not close to what is generally described as H. erectus s.s., whereas it appears, among H. erectus s.l., as an expression of the most distant morphology from this phenetic configuration (see for instance the MDS projection; Fig. 1c). At the same time, it shows affinities with H. ergaster samples and tends to cluster with them (see Fig. 1a,b). Daka should therefore best be seen as part of a local (African) evolutionary lineage spanning from approximately 1.8 Ma up to about 1 Ma. … The evidence provided here indicates a local phenetic continuity pattern, hence the occurrence in Africa of an anagenetic continuum from the emergence of the genus Homo up to about 1 Ma. Actually, Asfaw et al. (2002, p.318) do state that the “chronological and morphological seriation of the African fossils from KNM-ER 3733 / 3883 (Koobi Fora) to OH 9, to Daka / Buia to Bodo […] comprise a morphocline consistent with the hypothesis that they sample a single evolving lineage.” - Manzi et al (2003): link
Two key details from this excerpt are the following:
Homo ergaster, OH 9 and the Daka skull comprise a lineage that evolved in Africa
The Dmanisi specimens are temporally, geographically and morphologically intermediate relative to Homo ergaster and the classic Asian variation of Homo erectus (i.e. Peking Man & Java Man).
Therefore, given that the Dmanisi hominins have smaller brains than OH 9, the Daka skull and Asian Homo erectus specimens, they seem to have derived from an early wave of Homo erectus that migrated out of Africa; conversely, Asian Homo erectus seem to have derived from a later wave more related to OH 9 & the Daka skull. This implies that there were two waves of Homo erectus out of Africa, which aligns with the first scenario presented by Rogers et al: an initial expansion of Homo that occurred 1.9 Ma ago, and a second one that occurred between 0.9 and 1.4 Ma ago (which overlaps with the ages of OH 9 and the Daka skull). The initial expansion would have produced the primitive Dmanisi hominins, and the second expansion would have produced Asian Homo erectus.
This implies the following phylogeny:
Homo ergaster → African Homo erectus
African Homo erectus → Dmanisi Homo erectus
African Homo erectus → Later African Homo erectus
Later African Homo erectus → OH 9 & Daka skull
Later African Homo erectus → Asian Homo erectus
Below is a summary of conclusions that have been made herein and in articles that have been cited herein.
A superarchaic lineage contributed ancestry to Neandersovans and then exclusively to ancestors of Denisovans
The superarchaic lineage derived from an expansion of Homo out of Africa that occurred between 0.9 to 1.4 Ma ago
The superarchaic lineage was likely an expansion of Homo erectus
These conclusions are reflected in the following phylogeny, which builds upon the traditional phylogeny that was presented earlier herein:
Homo ergaster → African Homo erectus
African Homo erectus → Dmanisi Homo erectus
African Homo erectus → Later African Homo erectus
Later African Homo erectus→ OH 9 & Daka skull
Later African Homo erectus → Superarchaic Lineage
Superarchaic Lineage → Peking Man & Java Man
Superarchaic Lineage → Homo longi
Homo ergaster → Homo heidelbergensis
Homo heidelbergensis → Neandersovan
Neandersovan → Homo neanderthalensis & Asian Neandersovan
Asian Neandersovan + Homo longi → Denisovan
Homo heidelbergensis → Homo sapiens
This phylogeny is depicted in the graphic below. Note that superarchaic admixture in Neandersovans has been omitted, because it’s been concluded herein to be relevant to only the morphology of Denisovans (rather than Neanderthals); additionally, it seems to be negligible in Neanderthals as indicated by an article that will be referenced herein.

A graphic depicting the phylogeny that’s been deduced herein. It models Homo longi (represented by Yunxian 2) as a sibling lineage of Java Man & Peking Man
Note that Dmanisi Skull 5 (D4500-D2600) is depicted as an australopithecine rather than as Homo erectus, which is consistent with the previously-cited conclusions of Nery et al (2025); it’s depicted specifically as Australopithecus africanus, because its morphology seems to be most similar to this species (compare Dmanisi Skull 5 here to Australopithecus africanus Cranium Sts 5 here). Given that australopithecines and Homo ergaster would have been closely related groups with overlapping environmental niches, it’s not unreasonable for subgroups of each to have wandered out of Africa alongside one another ~1.9 Ma ago.
This phylogeny contradicts that suggested by Feng et al.
Our Bayesian tip-dating analysis also revealed that the diversification of Homo and the origin of H. sapiens have much greater time depths. The origin of the longi clade can be inferred to be about 1.2 Ma, probably slightly older than the Yunxian fossils. The origin of the sapiens clade is estimated to be about 1.02 Ma, also close to the age of Yunxian. The divergence between the longi clade and the sapiens clade is at about 1.32 Ma. The monophyletic Neanderthal clade, widely thought to be sister to H. sapiens, diverged from the longi and sapiens clades at about 1.38 Ma in our analysis.
This excerpt conveys Feng et al’s hypothesis that Homo longi and Homo sapiens comprise a clade to the exclusion of Neanderthals. However, as already shown herein, Roger et al’s genetic analyses indicate that Neanderthals and Denisovans - not Homo sapiens - carry admixture from a superarchaic lineage, which has been concluded herein to have produced Homo longi. Therefore, Neanderthals and Denisovans should comprise a clade with Homo longi to the exclusion of Homo sapiens. Furthermore, Roger et al further suggests that Neanderthals and Denisovans derived even their non-superarchaic ancestry from a common source (i.e. Neandersovans) - also to the exclusion of Homo sapiens. This is supported by the f-statistics that are shown below, which clearly show that Neanderthals and Denisovans are more closely related to each other than they are to Homo sapiens.

F2 statistics measure the genetic distance between two populations, whereby a smaller value equals a smaller genetic distance. These two bar graphs of f2 statistics clearly indicate that Denisovans and Neanderthals are closer to each other than they are to various Homo sapiens populations; they are based on calculations made via AdmixTools 2.0 that can be seen here.
The cladal relationship of Neanderthals and Denisovans is also supported by a recently released article by Population Geneticist David Reich: Hypothesis: A modern human range expansion ~300,000 years ago explains Neandertal origins (2026) [link]. Note that this article is pending peer-review.
Figure 1 of Reich’s article is shown below, alongside an edited version of it; the latter includes an additional label (“LCA”) and images of fossils to help visualize the morphological differences between the included populations. As can be seen, the original figure features a node named Eurasian archaic that leads to Sima de los Huesos and Denisovans.
Sima de los Huesos refers to hominins that were discovered at the eponymous archaeological site in Spain. These hominins have been estimated to have lived ~430 kya and to have been late members of Homo heidelbergensis, whose lineage evolved into Neanderthals; therefore, they were Proto-Neanderthals. That the Eurasian archaic node leads to only them and Denisovans indicates that Neanderthals and Denisovans comprised a clade to the exclusion of Homo sapiens, which contradicts Feng et al’s position.
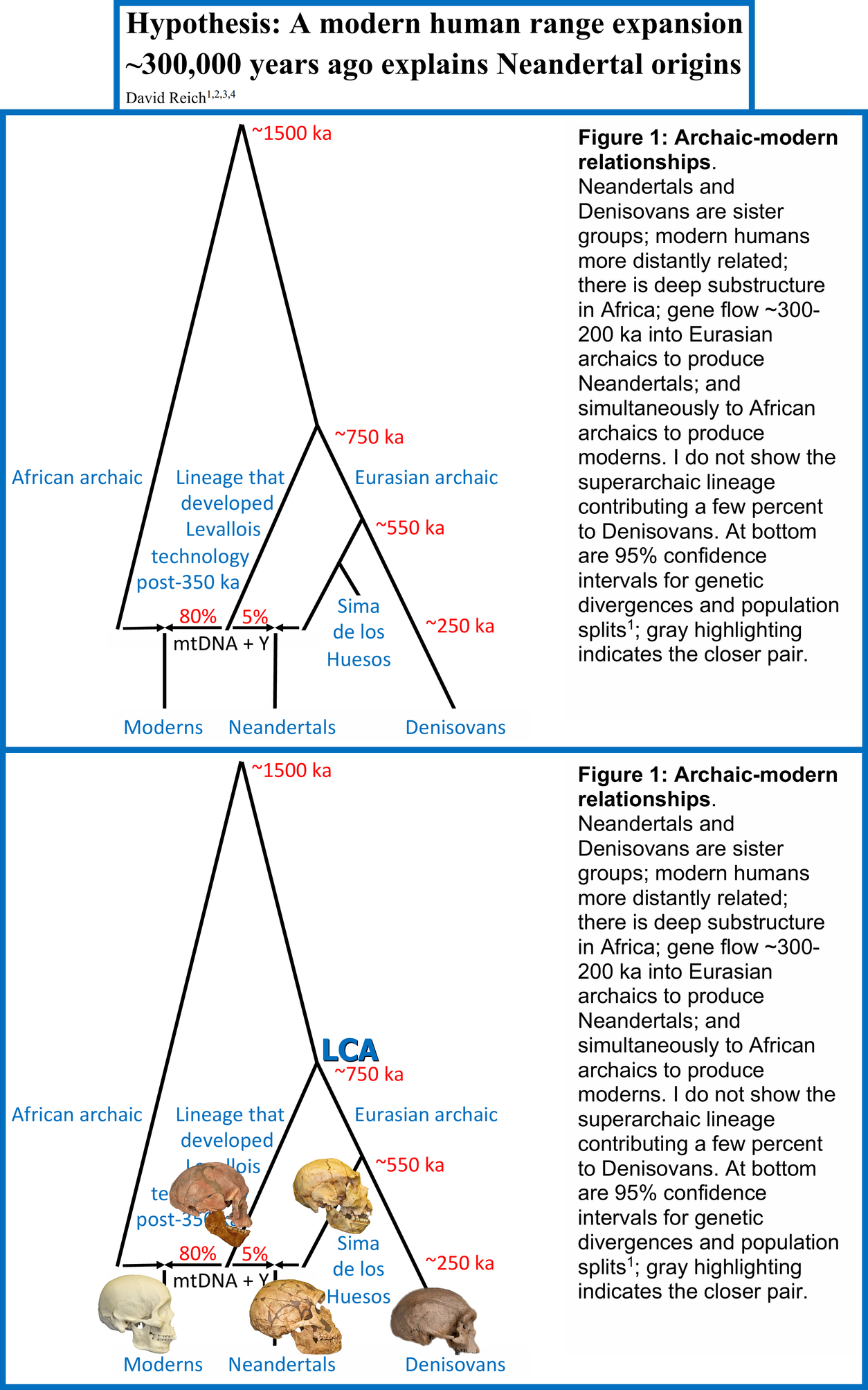
Figure 1 of Reich (2026)
Below are explanations of the nodes and populations that are featured in the figures.
LCA: the Last Common Ancestor of Homo sapiens (“Moderns”), Neanderthals (“Neandertals”) and Denisovans.
The ~750 ka Node: the point at which the LCA split into the Lineage that developed Levallois technology post-350 ka (Homo sapiens) and the Eurasian archaic lineage (Neandersovans)
Lineage that developed Levallois technology post-350 ka: the primary lineage of Homo sapiens, which provided 80% of the ancestry in Homo sapiens and 5% of the ancestry in Neanderthals.
African Archaic: an archaic lineage that split 1.5 million years ago from the lineage that produced the LCA. The figure indirectly indicates that this lineage provided 20% of the ancestry in Homo sapiens (“Moderns”), making it the secondary lineage of Homo sapiens.
Sima de los Huesos: Hominins that were discovered at the eponymous archaeological site in Spain. These hominins have been estimated to have lived ~430 kya and to have been late members of Homo heidelbergensis whose population was ancestral to Neanderthals. Therefore, they were Proto-Neanderthals.
Note that this figure does not model superarchaic admixture in Neanderthals or Denisovans; additionally, the supplementary text mentions its omission from the figure only in regard to Denisovans, which indicates that it’s likely negligible in Neanderthals. Despite its omission, Neanderthals and Denisovans are still modelled as having derived the majorities of their ancestries from a common source, to the exclusion of Homo sapiens.
Below is yet another edited version of Figure 1 from Reich (2026). It incorporates all of the conclusions that have been drawn herein regarding the hominin phylogeny.

Edited version of Figure 1 from Reich (2026)
As already stated herein, Denisovans’ morphological similarities to Yunxian 2 are likely the result of them carrying more superarchaic admixture than Neanderthals. This indicates that the morphology of Neanderthals may be less influenced by superarchaic admixture and therefore more similar to that of Neandersovans (“Eurasian archaic”).
The Homo heidelbergensis, Proto-Neanderthal specimens of Sima de los Huesos are the oldest specimens of the Neanderthal lineage, and they are modelled in Reich (2026) as having lacked Homo sapiens admixture - unlike proper Neanderthals; therefore, they should be the best representatives available for Neandersovan morphology. Furthermore, the best preserved specimen among them is Atapuerca 5 (link), which is a nearly complete skull.
As can be seen in the photos that are below, the overall cranial shape of Atapuerca 5 is quite similar to that of Turakana Boy from all angles. The differences between them primarily stem from the former being more robust; Atapuerca 5 has a larger brain case (1125 cc vs 880 cc), thicker brow ridges and larger zygomatics.

Composite of images featuring Atapuerca 5 & Turkana Boy
Interestingly, anthropological analyses indicate that Atapuerca 5 was an adult male and that Turkana boy was a pre-adolescent male. Therefore, given that Atapuerca 5’s morphology resembles that of Turkana Boy, Neandersovans appear to have been a variant of Homo heidelbergensis that neotenously evolved from Homo ergaster - neotenous meaning to retain immature traits in adulthood.
This seems to also apply to Homo sapiens, as can be seen in the below image.
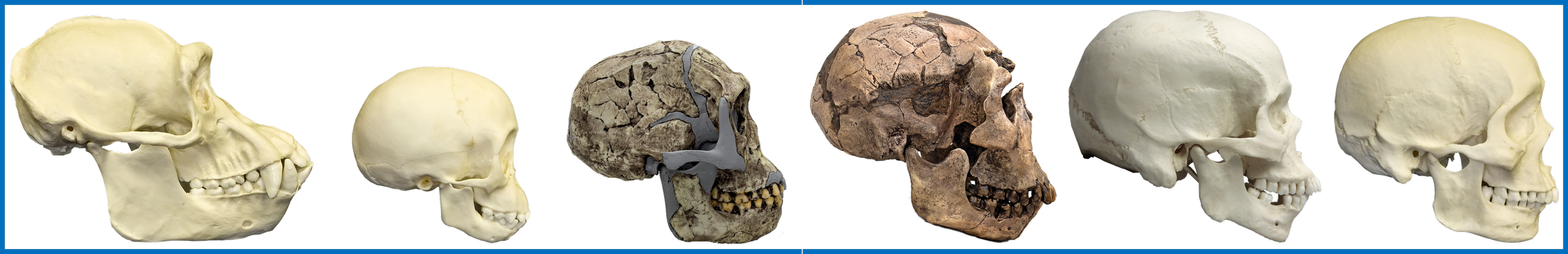
Neoteny is actually a key characteristic of the hominin lineage, which is why pre-adolescent and more mature Homo crania actually bear strong morphological similarity to those of infant chimpanzees (see the above image). This is consistent with the divergence between panins and hominins being the most recent divergence between extant ape lineages; the recency of this divergence indicates that the morphological traits that are shared by infant chimpanzees and more mature Homo individuals were present in our common ancestors.
Given that Reich (2026) models Homo sapiens’ primary source of ancestry as a sibling to Neandersovans, the two lineages must have derived their neoteny from the LCA. Therefore, it’s , and Neandersovans and proto-Homo-sapiens would have simply been subsets of it that subsequently exhibited the same neoteny.
_____________________________________
Share your agreements or disagreements below, in the comment section. Doing so can spark discussions through which alternative hypotheses or extensions of the one presented herein can be explored.